

Описание гаплогруппы I1-M253

Основной ареал рода I1 — северная Европа, с плавным понижением к югу. Наибольшие пики частот приходятся на Скандинавские страны. В южной Финляндии, и в некоторых регионах Швеции — около 45-50%, в Норвегии около 40%, у финских саамов линия I1 представлена 28%. В целом в Швеции и Финляндии частоты I1 имеют градиент от 15 до 45%. В Германии, в сновном, на севере страны частота I1 достигает 25%, в Дании — около 17%, а в Эстонии уже около 15%. Частоты порядка 12% зафиксированы во французской Нормандии. Также частота I1 высока на Британских островах, в особенности на побережье.
В центральной части Европы идет значительное снижение частоты. В Чехии, Словакии, Польше, Швейцарии и Венгрии — 5-7%. На Балканах: в Словении и в Македонии — 8-10%; в Сербии и Хорватии — около 5%; в Албании, Боснии, Греции — от 1 до 3%. На иберийском полуострове от 0 до 3%. В Молдавии 3%, а в Румынии около 2%. В Италии от 1 до 3%.
В восточной части Европы, например, на востоке Белоруссии до 7%, а к югу снижение до 3-4%. На Украине в среднем 4-5% линии I1. В России также наблюдается повышение частот на севере, и понижение на юге. Так, например, в Архангельской области зафиксировано до 14% представителей рода I1, в Вологодской — 11-12%, в Ивановской, Тамбовской и Пензенской областях — 10-11%, в Курской и Брянской областях — около 8%, а в Тверской — 5-7%. Еще один маскимум в Рязанской области - около 14%, и в приволжье 7-12%.
Карты гаплогруппы I1 в Европе и Скандинавии.

Мнение National Geographic о миграции гаплогруппы I1 (M253)

Карта Гаплогруппа I

Карта Гаплогруппа I1 (по старой классификации I1a)
Карта Гаплогруппа I1 (по старой классификации I1a)

Карта Гаплогруппа I1а (Субклад)



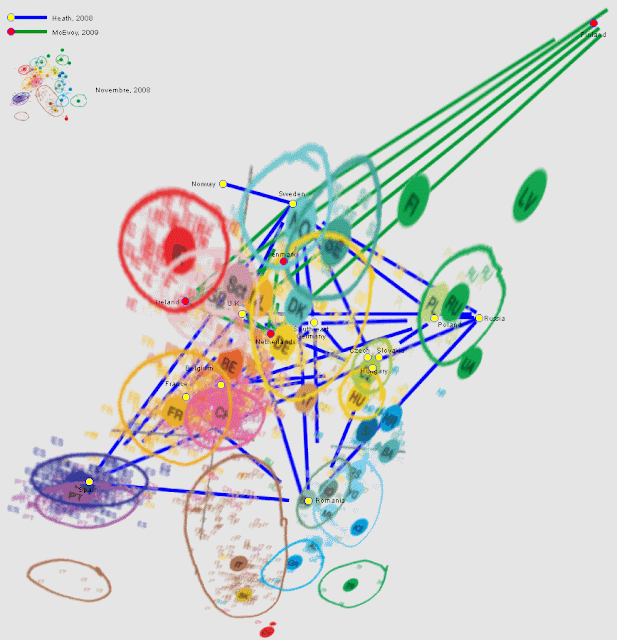
Dienekes Pontikos опубликовал результаты статистического анализа аутосомных SNP-маркёров 871 образцов из 60 популяций Западной Евразии. Полученное деление на кластеры выявило предсказуемую структуру, которая, тем не менее, заслуживает самого пристального внимания.Для тех, кто не хочет влезать глубоко в детали, сразу приведем структурную диаграмму.

http://drfaust-spb.livejournal.com/13731
МитоДНК и родство норвежцев с другими европейцами

Ради любопытства или для примера расмотрим ролик с чухонскими лицами у норвежцев.




Как видно, первая главная компонента отделяет Западную Евразию от Индии, вторая задаёт клинальный градиент между северными европейцами и средиземноморцами, третья выделяет из массива Западной Евразии североафриканское влияние, четвёртая — ашкеназов, пятая — финнов.Обращает на себя внимание плотная группировка русских со скандинавами, скандинавов — с немцами и ирландцами, половины немцев — с жителями Балкан, и относительная близость испанцев к северным европейцам. Показательно также, что по второму компоненту индусы находятся строго на уровне северных европейцев, не смещаясь по средиземноморской оси в сторону турок, армян и прочих греков.
Из книги
"Антропология Европы" (William Zebina Ripley)

























Генетическое разнообразие народов Европы.

Славяне и скандинавы наиболее однообразны, после британцев.
http://dienekes.blogspot.ru/2009/02/auton-et-al-on-global-distribution-of.html
ДНК древнего населения Скандинавии
Преемственность генофонда на севере Европы. Ниже речь пойдёт о результатах исследования ДНК древнего населения на территории Швеции, которое нашло ближайшее сходство среди современных латышей.
Краткая предыстория. Около 4000 гг. до н.э. на севере Центральной Европы формируется культура воронковидных кубков, представители которой живут плодами животноводства и земледетия. К 3300 годам до н.э. эта культура проникает в Южную Скандинавию, где на этот момент существует культура ямочной керамики, основу которой составляет охота, рыбная ловля и собирательство. Два подхода к хозяйствованию существуют в Скандинавии параллельно примерно до 2000 гг. до н.э., когда культура ямочной керамики полностью замещается земледельцами.
Перевод введения:
Уже больше ста лет идут дебаты о причине, побудившей древних жителей Европы сменить способ хозяйствования и перейти от охоты и собирательства к земледелию (неолитизация). Особый интерес представляет вопрос, изменился ли способ хозяйствования по причине смены населения или земледелие распространилось как результат культурного обмена.
В этой дискуссии Скандинавия занимает особое место, поскольку на её территории находился один из последних наиболее крупных культурных комплексов охотников-собирателей неолитической Европы — культура ямочной керамики. Любопытно, что эти поздние охотники и собиратели более тысячелетия жили параллельно с ранними земледельцами до момента своего исчезновения около 4000 лет назад. Длительное сосуществование двух типов культур в Скандинавии демонстрировалось в качестве аргумента против смены мезолитического населения современным.
Анализ образцов ДНК, извлечённых из останков древних жителей Скандинавии, показывает, чтопредставители культуры ямочной керамики не были прямыми предками современных скандинавов (включая саамов Северной Скандинавии) и имеют куда более близкое отношение к современному населению Восточной Балтики. Полученные нами результаты поддерживают гипотезу о смене неолитического населения Скандинавии на пост-неолитическое, восходящую к археологии. К тому же, наши данные согласуются с представлением о населении восточно-балтийского региона как потомках ряда популяций охотников и собирателей.
Helena Malmström et al., Ancient DNA Reveals Lack of Continuity between Neolithic Hunter-Gatherers and Contemporary Scandinavians, Current Biology, 24 September 2009.
Полная статья в формате PDF
Авторы исследовали 22 образца митохондриальной ДНК: 19 для представителей культуры ямочной керамики (захоронения на острове Готланд, 2800–2000 гг до н.э.) и 3 для представителей культуры воронковидных кубков (Gökhem, юго-запад Швеции, 3500–2500 гг. до н.э.).

Фиолетовый — ареал культуры ямочной керамики,
красная линия — граница распространения мегалитических сооружений
Проведённый анализ выявил кардинальное отличие структуры женских генофондов представителей двух культур (насколько о различии можно говорить на основе немногочисленных исходных данных): гаплотипы культуры ямочной керамики находят минимальное соответствие среди образцов культуры воронковидных кубков: из 19 образцов совпадает всего 2, около 11%.
Генетические расстояния от образцов ДНК культуры ямочной керамики до современных народов, от наиболее близких к самым далёким:
| народ | образцов | расстояние |
| латыши | 114 | 0,036 |
| литовцы | 163 | 0,040 |
| эстонцы | 117 | 0,047 |
| шведы | 289 | 0,052 |
| финны | 79 | 0,053 |
| норвежцы | 323 | 0,062 |
| саамы | 32 | 0,250 |
Нижняя часть списка отчёркнута как превышающая расстояние от древних ДНК до усреднённой современной европейской популяции, которое составляет 0,043.
Было бы любопытно взглянуть на расстояния от северо-западных русских и поморских поляков — они наверняка так же нашли бы большое сходство, но авторы этих данных не приводят. Основанием для этого предположения является тот факт, что значительная доля (8 из 19, 42%) образцов ДНК культуры ямочной керамики принадлежит к гаплотипу H1b, наибольшая доля которого констатируется среди восточных славян, эстонцев, на юго-западном побережье Балтики и Русском Севере [E.-L. Loogväli et al., 2004].
В любом случае, начинает проясняться резкое различие между населением Западной и Восточной Европы в пределе последних 5000 лет, которое говорит о нескольких путях заселения континента, а не примитивном расселении из одного центра, представление о котором бытовало раньше. Кроме того, саамов можно смело вычёркивать из предков современных европейцев.
Гаплогруппы в Щвеции

Анализируя краниологические серии с территории Европы эпох мезолита и неолита — [Pinhasi, von Cramon-Taubadel, 2011].Для восстановления картины этнических процессов в Европе данных одной антропологии недостаточно, поэтому для сравнения результатов следует кратко остановиться на данных другой области — популяционной генетики, а конкретно — маркёрах митохондриальной ДНК, которые передаются только по материнской линии. База древних образцов мтДНК на сегодняшний день превышает 1000 образцов и по своему объёму сопоставима с данными палеоантропологии.Чтобы прояснить направления переноса генетических маркёров, на основании сводногоперечня древних евразийских ДНК, на карты нанесена выборка из 346 образцов мтДНК мезолита, неолита, медного и бронзового веков. Цвет популяций для единства обозначений выбран в соответствии с упомянутым выше исследованием антропологии: красные — мезолит Европы, серые — неолит Малой Азии, синий — неолит Европы, зелёный — эпохи меди и бронзы. Образцы из евразийских степей не учитывались.
 |

Женская гаплогруппа U5 мтДНК - древнейшая в Европе.
Карта U5a:

Карта U5b:


Гаплогруппа U5a имеет возраст более 20 тыс. лет, не встречается в существенной частоте за пределами Европы, и на этом основании нередко именуется одной из древнейших европейских гаплогрупп. Гаплогруппа делится на две дочерние ветви, U5a1 и U5a2, возраст которых приближается к возрасту родителя, причем наибольшее разнообразие и максимум частоты обеих засвидетельствованы в центре и на востоке Европы. Обе ветви в умеренной частоте встречаются также за пределами Европы, в т.ч. в Азии, причем U5a1 еще в неолите проникла на территорию Южной Сибири, о чем можно судить по наличию ее обособленных вариантов у коренных народов востока России. О давнем проникновении европейцев в Восточную Сибирь свидетельствуют также результаты типирования древней ДНК: на стоянке Локомотив вблизи оз. Байкал, VI тыс. до н.э., обнаружен образец, принадлежащий к U5a. Частоты U5a на Востоке Европы в целом следуют общеевропейским закономерностям в распространении этой гаплогруппы: на севере ее средние частоты возрастают в 1.5-2 раза по сравнению с частотами на юге, однако разнообразие остается умеренным и достигает своего максимума в центре ареала. Максимум частоты U5a найден у сету, марийцев, чувашей а также некоторых народов Северного Кавказа.
Результаты исследований палеоДНК:| Гаплогруппа | Возраст | N | Арх. культура, регион | Источник |
| U5a1 | 9-10 тыс. лет | 1 | Лебяжинка, Россия | Bramanti et al. 2009 |
| U5a1a | Неолит | 1 | Derenburg, Meerenstieg II (LBK culture) | Haak et al. 2011 |
| U5a | 10 тыс. лет | 1 | Чекалино, Россия | Bramanti et al. 2009 |
| U5a | 8,700 лет | 1 | Hohlenstein-Stadel, Германия | Bramanti et al. 2009 |
| U5a | 6 тыс. лет | 1 | «Чедерский человек», Англия | Bryan Sykes 2002 |
| U5a | 4,250 лет | 1 | Drestwo, Польша | Bramanti et al. 2009 |
| U5a1 | 3-4,300 лет | 2 | Андроновская культура, Россия | Keyser et al. 2009 |
| U5a1a | 2 тыс. лет | 1 | Монголия | - |
| U5a1 | 13-12 в. до н.э. | 1 | Izmaylovka (Kourgan 7,10), Казахстан | Lalueza-Fox et al. 2004 |
| U5a1a | 1,200 до н.э. | 1 | Grave Circle B (Mycenae), Греция | Abigail et al. 2009 |
| U5a | Неолит | 2 | Fridtorp, Европа | Malmström et al. 2009 |
| U5a1 | 10-11 в. | 1 | Регион Карпат, Венгрия | Nagy et al. (2011) |
| U5a | 12-13 в. | 1 | Apollo Klarios (Византия) | Ottoni et al. (2011) |
Гаплогруппа U5b

Гаплогруппа U5b является одной из древнейших линий Западной Евразии. В целом U5b нельзя отнести в числу частых линий: она распространена в Европе и на северо-западе Африки со средней частотой 3%-6% и большинство известных случаев повышения ее частот у той или иной популяции объяснимо дрейфом. За пределами указанного ареала, например на Ближнем Востоке, в Центральной Азии и Сибири средние значения частот ниже U5b 1%. U5b - единственная из европейских гаплогрупп, обнаруженных в значительной частоте у негроидных популяций Западной Африки. Средняя частота U5b в Европе составляет 4%. Наиболее примечательное исключение - саамы, у которых данная гаплогруппа представлена лишь одной ветвью U5b1b1a, но почти у половины популяции. Также высока частота U5b у карел (13%), мордвы (11%) и коми-зырян (7%). Другой локальный максимум обнаруживается на юге Центральной и Восточной Европы: у словаков, украинцев, венгров и белорусов средняя частота U5b составляет около 5% в то время как среднерусская частота менее 3%.
Результаты исследований палеоДНК:| Гаплогруппа | Возраст | N | Арх. культура, регион | Источник |
| U5b1c | 18,000-15,000 до н.э. | 1 | Nerja, Málaga (Солютер), Испания | Fernández et al. (2005) |
| U5b2 | 7,200 до н.э. | 1 | Falkensteiner, Германия | Bramanti et al. (2009) |
| U5b2 | 6,700 до н.э. | 1 | Hohlenstein-Stadel, Германия | Bramanti et al. (2009) |
| U5b2 | 6,300 до н.э. | 1 | Donkalnis (Kunda Culture), Литва | Bramanti et al. (2009) |
| U5b2 | 4,450 до н.э. | 1 | Kretuonas (Narva Culture), Литва | Bramanti et al. (2009) |
| U5b2 | 4,200 до н.э. | 1 | Kretuonas (Narva Culture), Литва | Bramanti et al. (2009) |
| U5b1 | 3,650 до н.э. | 2 | Dudka Culture, Польша | Bramanti et al. (2009) |
| U5b1c | 3,000 до н.э. | 1/29 | Treilles, Франция | Lacan et al. (2011) |
| U5b | 2,600 до н.э. | 1 | Eulau, Германия | Haak et al. (2008) |
Гаплогруппа R1a в Скандинавии
21.5.2. Скандинавия
Гаплотипы этой ветви типичны для жителей Скандинавского региона (21 из 56 гаплотипов в Норвегии, Швеции и Дании, т.е. 38%), но её география не ограничивается только им. Одна из подветвей (жёлтые маркеры на карте) захватывает Центральную Европу. Кроме того, родословная многих англичан и шотландцев также восходит к этой ветви. Скандинавские гаплотипы найдены также в Финляндии, как среди этнических шведов, так и финнов.
Распредение скандинавских ветвей гаплотипов показано на рис. 21.5.2.1.

Скандинавская группа гаплотипов отличается тем, что большинство их имеют в предковом гаплотипе DYS19 = 15, DYS3891 = 14, DYS447 = 23, и пару YCAII = 19-21, в то время как в большинстве европейских и евразийских предковых гаплотипах там соответственно 16, 13, 24, и 19 - 23. Разница в три мутации между 25-маркерными европейскими предковыми гаплотипами помещает их общих предков примерно на 4200 - 4700 лет назад, так что ничего необычного и особенно древнего в этом нет. Таким образом, общие предки скандинавских гаплотипов гаплогруппы R1a1 те же самые, что общие предки большинства европейских ДНК-генеалогических ветвей.
Анализ 67-, 37- и 25-маркерных скандинавских гаплотипов показал, что их аллели на первых двух панелях полностью воспроизводятся для гаплотипов разной протяженности (третья панель по определению есть только у 67-маркерных гаплотипов). 67-маркерный базовый гаплотип для всего дерева, определенный по данным рис. 21.5.2.2, имеет следующий вид:
13 25 15 11 11 14 12 12 10 14 11 31 -15 9 10 11 11 23 14 20 32 12 15 15 16-11 12 19 21 16 16 17 18 34 38 12 11-11 8 17 17 8 12 10 8 11 10 12 22 22 15 11 12 12 13 8 14 23 21 12 12 11 13 11 11 12 12
На дереве 67-маркерных гаплотипов несколько выделяется ветвь из 11 гаплотипов справа внизу, состоящая из двух подветвей, из семи и четырёх гаплотипов. Почти все они имеют YCAII = 19,23.

Их базовые 25-маркерные гаплотипы соответственно, выглядят следующим образом:
13 25 15 11 11 14 12 12 11 14 11 31 - 15 9 10 11 11 24 14 20 31 12 15 15 16
13 25 15 11 11 14 12 12 10 13 11 30 - 15 9 9/10 11 11 23 14 20 31 12 13 15 15/16
Ветвь из четырех гаплотипов содержит 18 мутаций, что дает 2725±700 лет до общего предка. Ветвь из семи гаплотипов - 30 мутаций, что дает 2600±540 лет до общего» предка Но между ними - 4.7 мутаций, что помещает их общего предка примерно на 4100 лет назад. Это предок имел типичную для Европы пару YCA = 19, 23 и DYS565 = 13 (выделены), и его гаплотип был
13 25 15 11 11 14 12 12 10 13 11 30 - 15 9 10 11 11 24 14 20 32 12 15 15 16 -11 12 19 23 16 16 18 19 35 38 13 11 -11 817 17 8 12 10 8 11 10 12 22 22 15 10 12 12 13 8 13 23 21 12 12 11 13 11 11 12 13
Мы будем далее называть эту двойную ветвь «старой скандинавской ветвью».

Рис. 21.5.2.3. Дерево из 50 37-маркерных гаплотипов скандинавского клана (Рожанский и Клёсов, 2009).
Остальные три ветви на 67-маркерном дереве имеют следующие базовые гаплотипы и «возрасты» общих предков: (а) Ветвь из 7 гаплотипов:
13 25 15 11 11 14 12 12 10 14 11 31-15 9 10 11 11 23 14 20 32 12 15 15 16 - 11 12 19 21 16 15 18 17 35 39 12 11 -11 8 17 17 8 12 10 8 11 10 12 22 22 15 11 12 12 13 8 14 23 21 12 12 11 13 11 11 12 12
15 мутаций на семь 25-маркерных гаплотипов указывает на 1250±350 лет до общего предка (примерно 8-й век нашей эры, плюс-минус три-четыре столетия).
(б) Ветвь из 8 гаплотипов:
13 25 15 11 11 14 12 12 10 14 11 31 -15 8 10 11 11 23 14 20 31 12 15 15 16-11 12 19 21 16 16 17 18 34 38 12 11 - 11 8 17 17 8 12 10 8 11 10 12 22 22 15 11 12 12 13 8 14 23 21 12 12 11 13 11 11 12 12
24 мутации на восемь 25-маркерных гаплотипов указывает на 1750±400 лет до общего предка (примерно 3-й век нашей эры, плюс-минус четыре столетия).
(в) Ветвь из 11 гаплотипов:
13 25 15 11 11 14 12 12 10 14 11 31 - 15 9 10 11 11 23 14 20 32 12 15 15 16 - 11 12 19 21 16 16 17 18 34 37 12 11 - 11 8 17 17 8 12 10 8 11 10 12 22 22 15 11 12 12 13 8 14 23 21 12 12 11 13 11 11 12 12
32 мутации на 11 25-маркерных гаплотипов указывает на 1700±350 лет до общего предка (примерно 4-й век нашей эры, плюс-минус три-четыре столетия).
Как видно, все три «молодые» ветви очень похожи - все имеют общих предков в середине 1-го тысячелетия нашей эры, все имеют характерную пару YCAII = 19,21 и характерную аллель DYS565 = 12 в базовых гаплотипах. По историческим сведениям, именно в середине 1-го тысячелетия нашей эры в Скандинавию прибыли племена из Азии - из Прикаспия, Причерноморья или Приуралья, детали этого историкам неизвестны.
Все три 67-маркерных базовых гаплотипа различаются всего на 6.7 мутаций на 201 маркера. Это помещает их общего предка менее чем на 400 лет ниже усреднённых величин жизни этих трёх общих предков племён, то есть примерно на 1900 лет назад, во 2-й век нашей эры плюc-минус несколько веков.
Либо люди из всех трёх племён прибыли в Скандинавию с востока почти одновременно, либо разделение на ветви произошло уже в Скандинавии. В любом случае, эти племена очень близки по гаплотипам. Можно вспомнить норвежские саги, что Один пришёл с востока (из Асгарда) в Скандинавию с тремя сыновьями, но это будет уже на грани фантазий. Хотя и не исключено.
Предковый гаплотип комбинированной «молодой скандинавской ветви» следующий:
13 25 15 11 11 14 12 12 10 14 11 31 - 15 9 10 11 11 23 1420 32 12 15 15 16 - 11 12 19 21 16 16 17 18 34 38 12 11 - 11 8 17 17 8 12 10 8 11 10 12 22 22 15 11 12 12 13 8 14 23 21 12 12 11 13 11 11 12 12
Картина в целом подтверждается при рассмотрении дерева 25-маркерных гаплотипов (рис. 21.5.2.4). Видно, что это дерево расходится на две основные части - слева «молодая» ветвь, справа - «старая». При этом старая ветвь опять состоит из подветвей. Если их не учитывать, то все 26 гаплотипов этой старой ветви содержат 96 мутаций, что дает возраст общего предка 2200±310 лет. Это - уже знакомая нам усредненная цифра, полученная выше без учёта ветвей. Но «молодая» ветвь слева содержит 144 мутации на 49 гаплотипов, что даёт 1700±220 лет до общего предка. Это 4±2 век нашей эры. Время Великого переселения народов. Базовый 25-маркерный гаплотип этой ветви
13 25 15 11 11 14 12 12 10 14 11 31 -15 9 10 11 11 23 14 20 3212 15 15 16
Как уже отмечалось выше, подавляющее количество гаплотипов этой молодой ветви, пришедшей в Скандинавию в середине 1-го тысячелетия н.э., содержит пару YCAII = 1921, которой почти нет в старой ветви.
Эта левая «молодая» ветвь содержит свои подветви, и в качестве примера можно рассмотреть подветвь из 11 гаплотипов, между номерами 3 и 846. Она содержит 28 мутаций, что даёт 1475±320 лет до общего предка. Это опять середина 1-го тысячелетия, примерно 6-й век н.э. Базовый 25-маркерный гаплотип этой подветви
13 25 15 11 11 14 12 12 10 14 11 31 -15 8 10 11 11 23 14 20 32 12 15 15 16
что почти полностью совпадает с приведённой выше, отличись лишь на 0.75 мутации.
Итак, в качестве рабочей гипотезы можно предположить, что общий предок ветви R1a1, пришедший в Скандинавию в середине 1-го тысячелетия, прибыл из «Азии», как тогда называли причерноморские и прикаспийские степи, как и всё, что находилось за Доном. За три тысячи лет до того его предки мигрировали из Европы на Русскую равнину и ее южную, степную часть. В середине 1-го тысячелетия, во времена Великого переселения, потомки R1a1 вернулись обратно в Европу, неся дополнительные мутации в своих гаплотипах, и, в частности, мутацию 19-21 в YCAII, мутацию DYS3891=14, мутацию DYS447 = 23, DYS565=12 и ряд других мутаций.
Об этом прибытии «азиатских» племён в Скандинавию говорят многочисленные исторические сведения. Но перед рассмотрением этих представлений целесообразно рассмотреть, когда в Скандинавии появились люди. Обычно под Скандинавией - в этно-территориальном отношении - понимают собственно Скандинавский полуостров, северное побережье Балтийского моря, и современные Данию и Исландию. Заселение Исландии происходило только в 9-м веке н.э., поэтому в настоящей работе мы её рассматривать не будем. Хотя тот факт, что общий предок носителей гаплогруппы R1a1 Исландии жил примерно 4500 лет назад, является хорошим примером, что датировки общих предков серий гаплотипов нельзя автоматически переносить на датировку заселения территорий. Необходимо привлекать другие независимые сведения, если таковые доступны.
Монография «История Швеции» (М., 1974, стр. 51; цитируется по Ю.Н. Дроздову, 2008) сообщает, что «в конце среднего неолита, около 2-го тысячелетия до н.э., на юге Скандинавии появились кочевые скотоводческие племена, оставившие прекрасно отполированные орудия - боевые или ладьевидные топоры. Пришельцы оттеснили прежних обитателей страны и быстро освоили внутренние области Южной и Средней Швеции».
В качестве «прежних обитателей страны» вполне могли быть носители гаплогруппы 12, которые имеют общих предков в Европе - по данным ДНК-генеалогии - не менее 15 тысяч лет назад (см. выше). Другие кандидаты (в отношении столь древней гаплогруппы в Скандинавии) просматриваются плохо. Носители гаплогруппы 12 и могли относиться к культуре воронковидных кубков, в период примерно 5500 - 4500 лет назад.

Рис. 21.5.2.4. Дерево из 75 25-маркерных гаплотипов скандинавского клана. Из работы [Рожанский и Клёсов, 2009]. |
Тогда «кочевые скотоводческие племена» могли представлять культуру шнуровой керамики, иметь в основном гаплогруппу R1a1 и активно действовать в период 5000 -4000 лет назад. Их в значительной степени потеснила культура боевых топоров, с предполагаемыми носителями гаплогруппы R1b1, в период 4000 - 3000 лет назад. Это были опять же кочевые, скотоводческие племена, прибывшие, вероятно, из регионов курганной культуры. Естественно, эти культуры пересекались, сосуществовали в той или иной степени на разных территориях, и никогда не вытесняли полностью одна другую.
Такой порядок культур определяется тем, что по данным ДНК-генеалогии носители R1b только начинают появляться в Европе около 4000 лет назад, и, возможно, появились там в ряде регионов только 3600 лет назад [Клёсов, 2008f, Klyosov, 2009a,b; Клёсов, 2009а]. В примечании к академическому изданию Тацита («О происхождении германцев и местоположении Германии») отмечается, что «по современным научным воззрениям, Гaллия была заселена кельтами, двигавшимися с востока; около 2000 г. до н.э. кельтские поселения существовали на юге и юго-западе нынешней Германии».
До прихода с востока новой волны R1a1 в середине 1-го тысячелетия на юге Скандинавии и на севере Европы жили разнообразные племена. Видимо, это были носители гаплогрупп и R1a1, и R1b1, и I1, и I2. Другие гаплогруппы если и были, то определённо в меньшинстве. У Корнелия Тацита в его произведении «О происхождении германцев и местоположении Германии» (1-й век н.э.) упоминаются марсы, гамбривии, свебы, вандилии, тунгры и указывается, что это имена подлинные и древние.
Тацит указывает, что вместе с умершим у германцев огню предается иногда конь. Это принципиально отличается от восточного способа захоронения коня с хозяином в одной могиле, что начинает повторяться в Скандинавии с середины 1-го тысячелетия.
Племя свебов, которое по созвучию можно посчитать скандинавским, на самом деле германское, и по описанию Плиния Старшего («Естественная история», IV, 99 - 101) из пяти групп германских племён четвертая группа - гермионы, «живущие внутри страны», и в неё входили свебы, гермундуры, хатты и херуски. Видимо, свебы - это швабы, поскольку центральную часть Германии Тацит называет Свебией.
Далее, описывая племена к северо-западу от Германии, включая полуостров Ютландию, Тацит упоминает хавков. хаттов, фосов, фризов, херусков, кимвров. Описывая свебов, Тацит отмечает, что они расчленяются на много отдельных народностей, носящих свои наименования (например, семионы), хотя общее название их - свебы. Там же Тацит упоминает такие племена, как лангобарды, ревдигны, авионы, англии, варины, эвдосы, свардоны, нуитоны. Свебов Тацит упоминает и в связи с островом с предполагаемым современным названием Зеландия, на котором ныне находится Копенгаген. Стоит отметить, что языки древних германцев, как и британцев, описываются как индоевропейские в академическом примечании к произведению Тацита. Это опять даёт основания предполагать, что речь идет в основном о племенах гаплогруппы R1a1.
Одно из племён под названием наганарвалы поклонялись богам Кастору и Поллуксу. Интересно, что Кастор и Поллукс - божества ариев, многократно описанные в индийских Ведах, и древняя история почитания этих божеств уходит у ариев на 7000 лет назад, как установлено из астрономических данных по динамике появления этих звёзд на небосклоне [Клёсов, 2008а].
Наконец, Тацит упоминает племя свионов, живущих «посреди самого Океана», то есть в Скандинавии, поскольку Океаном Тацит называл Балтийское и Северное моря, а Скандинавию считал островом. За свионами, по описаниям Тацита, находился Северный Ледовитый океан. Свионов Тацит описывает как умелых мореплавателей.
Все эти описания Тацита относятся к началу нашей эры, до эпохи Великого переселения народов и прибытия новой волны носителей R1a1 в Скандинавию. По мнению Давида Фокса ([David Faux, 2007], препринт), эта миграционная волна практически не нашла отражения в научной литературе, поскольку крах Римской империи на века, фактически, вплоть до 13-го века (по другим данным, до 11 -12 вв.) остановил исторические исследования в Европе, и в особенности в Скандинавии.
Фокс связывает прибытие R1a1 с Кавказа и/или Средней Азии, хотя не даёт чётких обоснований такому отнесению. Фокс также полагает, что эта волна из Азии и породила викингов, этнически и генеалогически. Историю викингов часто отсчитывают с 793 года, с нападения на Линдесфарнский монастырь в Нортумбрии, Англия.
С другой стороны, Фокс идёт до крайностей, называя гаплогруппу R1a1 «редкой», и относя практически всех R1a1 в Европе к этой волне из Азии в середине 1-го тысячелетия.
Про археологические находки R1a1 в Европе, относящиеся к временами 4600 лет назад, Фокс тогда не знал. Фокс датирует миграцию R1a1 в Скандинавию с востока 375 - 550 гг. н.э. В то время в раскопках стали активно появляться фигурки с азиатскими мотивами, которые часто приписывают скифам. Фокс считает скифов племенем южной части Средней Азии, отсюда и его отнесение исходного региона миграции R1a1 в Скандинавию в середине 1-го тысячелетия н.э. Стиль и характер религиозных обрядов также резко изменился в Скандинавии в середине 1-го тысячелетия н.э.
Мы не будем здесь анализировать народные легенды, предания, саги, предположительно описывающие те времена. Это достаточно подробно описано в упомянутом препринте Фокса и в книге Ю.Н. Дроздова (2008). Коротко перечислим лишь несколько произведений. В «Саге о Скьёльдунгах» неизвестного автора говорится как Один, отец Скьёлда, придя из Азии и подчинив местных жителей, стал властвовать над северной частью Европы (Саксонией, Данией, Швецией).
При этом Один вышел из земель к северу от Меотийского болота, то есть территорий от Азовско-Прикаспийских степей до побережья Ледовитого океана, по представлениям античных и раннесредневековых авторов. Эти земли древние скандинавы называли «Свитьод Великий и Холодный». Интересно, что те времена описываются в «Саге об Инглингах» как «в те времена римские хёдвинги ходили походами по всей земле и подчиняли себе все народы», и тоже представляется переселенческая версия происхождения значительной части скандинавских народов.
Об этом же повествует «Описание Земли III» в историко-географическом сочинении 13-го века - «Север заселили тюрки и люди из Азии. Поэтому с уверенностью можно сказать, что вместе с ними пришел сюда на север и язык, который мы называем северным, и распространился этот язык по Саксланду, Данмору и Свитьод, Норвегии и по некоторой части Энгланда. Главой этого народа был Один, сын Тора». Напоминаем, что территории восточнее Танаиса (Дона) античные источники называли Азией.
Подводя итог настоящему разделу, укажем, что скандинавские гаплотипы гаплогруппы R1a1 принадлежат двум группам, в отношении их общих предков. Древняя группа имеет общего предка, жившего примерно 4100 лет назад, что находится в хорошем соответствии с археологическими данными, предположительно культура шнуровой керамики. Относительно молодая группа, или ветвь, имеет общего предка 1700±220 лет назад (примерно 4-й век н.э.) или 1475±320 лет назад (примерно 6-й век н.э.). Это тоже хорошо согласуется с археологическими и историческими данными, хотя они стали очерчиваться лишь недавно.
14.1. Скандинавия (Дания, Швеция, Норвегия, Финляндия)
Выборка, полученная в базе данных YSearch, состоит из 68 25-маркерных гаплотипов (рис. 14.1.1), которые расходятся по двум разным ветвям, «старой» (на дереве слева) и «молодой» (на дереве справа).

Рис. 14.1.1. Дерево 25-маркерных скандинавских гаплотипов гаплогруппы I2-M438, построено по данным базы данных Ysearch. Дерево состоит из 68 гаплотипов. Индексы гаплотипов соответствуют Швеции, Норвегии, Дании, Финляндии.
В свою очередь, «старая ветвь» расходится на несколько ветвей (рис. 14.1.2).

Рис. 14.1.2. Дерево 25-маркерных скандинавских гаплотипов «старой ветви» гаплогруппы I2-M438 (слева на рис. 14.1.1). Дерево состоит из 33 гаплотипов. Индексы гаплотипов соответствуют Швеции, Норвегии, Дании, Финляндии.
Рассмотрим ветви по порядку. 14 гаплотипов на рис. 14.1.2 на ветви справа содержат 69 мутацию от базового гаплотипа
13 22 14 10 13 14 11 14 11 12 11 28- 15 898 11 23 16 20 281214 15 15
что даёт время жизни общего предка ветви 3025±470 лет назад.
19 гаплотипов левой ветви содержат 242 мутации от базового гаплотипа
13 23 14 10 14 15 11 14 11 12 11 28 - 16 8 9 8 11 24 16 20 28/2912 14 14 15
что даёт время жизни общего предка ветви 9575±1140 лет назад.
Между этими двумя ветвями почти восемь мутаций (выделены), что помещает их общего предка примерно на 9000 лет назад. Таким образом, правая ветвь - дочерняя левой. Левая ветвь и есть предковая для «старой ветви» скандинавских гаплотипов гаплогруппы I2, они же левая ветвь на рис. 14.1.1.

Рис. 14.1.3. Дерево 25-маркерных скандинавских гаплотипов «молодой ветви» гаплогруппы I2-M438 (справа на рис. 14.1.1). Дерево состоит из 35 гаплотипов. Индексы гаплотипов соответствуют Швеции, Норвегии, Дании, Финляндии.
Правая, «молодая» ветвь на рис. 14.1.1 в свою очередь тоже разделяется на несколько подветвей, из которых совершенно отдельно стоит только самая молодая ветвь слева вверху (рис. 14.1.3)
Базовый гаплотип этой ветви 14 23 15 10 15 15 11 13 11 14 12 31 - 15 9 10 11 11 25 14 20 27 12 14 14 15
от которого все семь гаплотипов ветви имеют 28 мутаций. Это даёт 2425±520 лет до общего предка.
Остальная часть дерева из 28 гаплотипов имеет всего два отличия в мутациях (отмечено) от базового гаплотипа «молодой» ветви, и содержит 170 мутаций от базового гаплотипа
14 23 15 10 15 15 11 13 11 14 12 31 - 15 8 10 11 11 25 14 20 27 11 14 14 15
Это даёт 3850±490 лет до общего предка ветви из 29 гаплотипов и помещает общего предка всего дерева в тот же временной интервал. Малая ветвь из семи гаплотипов оказалась дочерней.
Итак, на общем дереве скандинавских гаплотипов гаплогруппы I2 имеются всего две основные ветви, «возрастом» 9575±1140 лет и 3850±490 лет, с базовыми гаплотипами
13 23 14 10 14 15 11 14 11 12 11 28 - 16 8 9 8 11 24 16 20 28/29 12 14 14 15
14 23 15 10 15 15 11 13 11 14 12 31 - 15 8 10 11 11 25 14 20 27 11 14 14 15
соответственно. Между ними - 18.5 мутаций в 25-маркерных гаплотипах, что помещает общего предка гаплотипов I2 в Скандинавии примерно на 14925 лет назад.
Структура генофонда Швеции
Заботливые финские генетики опубликовали статью с результатами исследования генофонда своих соседей, шведов, и его сравнения с генофондами соседних народов севера Европы.
Ниже перевод фрагментов статьи, много диаграмм, много букв и ряд любопытных соображений, на которые наводит это исследование.

В исследовании использовались аутосомные аллели однонуклеотидного полиморфизма (SNP) со всей длины генома, которые наследуются от обоих родителей, в отличие от однородительских Y-ДНК и мтДНК. Выборка включала в общей сложности 379000 SNP, принадлежащих 1525 шведам из разных частей страны и 3212 представителям других народов: финнам (запад и восток), северным немцам (Киль), британцам (несколько регионов) и северным русским (Вологда). Наличие данных о популяциях каждого административного субъекта Швеции и Финляндии позволило в точности проследить географическую изменчивость шведского генофонда.
Swedish Population Substructure Revealed by Genome-Wide
Single Nucleotide Polymorphism Data
Elina Salmela et al.
Abstract
Генетические образцы шведов — особенно из южных регионов Швеции — оказались близки немцам и британцам, в то время как генетическое расстояние до финнов оказалось существенно больше. Общая структура генофонда Швеции показала клинальную изменчивость, а переходная область в южных и центральных регионах оказалась едва уловимой. Напротив, северная часть Швеции — Norrland — показала явное отличие генофонда как от других регионов страны, так и от соседних народов. Основными причинами такого разительного отличия, возможно, стали значительная удалённость этого региона и генетический дрейф вследствие её слабой заселённости. Внутреннее генетическое разнообразие на территории Швеции оказалось более заметным, чем внутри многих других популяций Центральной Европы, хотя и меньше обнаруженного, например, в Финляндии.
Статья полностью: PLoS ONE 6(2): e16747. doi:10.1371/journal.pone.0016747
1. Структура Швеции
Проведённый авторами статистический анализ выявил неоднородную структуру населения Швеции, границы компонентов которой примерно соответствуют трём историческим регионам: Götaland, Svealand и Norrland.

В целом, для территории Швеции характерна изменчивость генофонда с юга на север. Как показывают результаты многомерного шкалирования (MDS), два южных региона Götaland и Svealand показывают значительное сходство между собой и формируют очень плотный кластер, в то время как население региона Norrland от них заметно отличается. Степень различия видна из диаграммы MDS (C), где образцы из Norrland (зелёные) формируют отдельный кластер от Götaland и Svealand. В то же время, внутри региона Norrland заметное различие по второй оси показывают две северных области: Västerbotten (VBO) и Norrbotten (NBO).

По частоте аллелей Svealand и Götaland оказались очень близки, а различия между ними и северным регионом Norrland оказались сопоставимы с расстоянием между Германией и Британией. По уровню межпопуляционной изменчивости (Fst) население Швеции оказалось более разнообразным, чем население Германии и Британии, уступая различиям внутри Финляндии. Генетическое разнообразие в Norrland также оказалось более значительным, чем в других регионах Швеции.
Такая заметная обособленность севера Швеции дала авторам повод более пристально рассмотреть регион Norrland:
На диаграмме D отмечены образцы из провинции Norrland, обозначенные различными цветами с юга на север, в соответствии с расположением долин 10 крупнейших рек. На диаграмме заметно ясное деление населения региона на 3 кластера: северный, центральный и южный. Заслуживает внимания тот факт, что центральный кластер отличается только по первой оси, а северный — исключительно по второй.
Попарное сравнение всех областей страны по индексу структурного сходства (IBS) показало идентичный с MDS (диаграмма D) результат: две северных области региона Norrland — Västerbotten (VBO) и Norrbotten (NBO) — заметно отличаются как от остальной территории Швеции, так и друг от друга.
Такой расклад навёл авторов на предположения о причинах разительного отличия населения региона Norrland:
Северная часть Швеции — Norrland — показала ясно выраженную структуру населения, которая могла бы объясниться дрейфом генов в слабо населённом регионе. Однако, эта гипотеза выглядит сомнительной в связи с тем, что в регионе не наблюдается заниженного генетического разнообразия — фактически, Norrland показал внутри себя значительно меньшее структурное сходство, чем Götaland. Всё это указывает на существование в регионе Norrland изолированных популяций, что подтверждается результатами многомерного шкалирования и межпопуляционной изменчивости.
С другой стороны, выраженная структура и высокое разнообразие отчасти могут являться результатом значительного притока инородного населения. Проблема в том, что влияние со стороны финнов является умеренным. Возможно, причина заключается во влиянии саамских или норвежских компонентов, сравнение с которыми нами не проводилось.
Короче говоря, на севере страны обнаружились особые шведы, которые отличаются от остальных шведов настолько же, насколько немцы от британцев. Причины может быть две. Первая: это население живёт на севере Швеции с очень давнего времени — по крайней мере, с более раннего, чем прибытие из континентальной Европы предков современных южных шведов, — и сохранило с тех пор свою разнообразную структуру. Вторая: в Norrland намешано столько разнородных компонентов, что регион в целом стал заметно отличаться от всех своих соседей. При всём этом, наличие инородных компонентов не отрицает возможности наличия древнего аборигенного населения, поскольку сам Norrland делится на три отчётливых кластера — NBO, VBO и остальные области.
Для прояснения процессов сложения структуры населения необходимо взглянуть на результаты сравнения шведов с соседними народами.
2. Сравнение Швеции и Финляндии
В целом, генетическое расстояние между северными шведами и восточными финнами значительно превышает географическое, что говорит о слабом влиянии финнов на население Швеции. Уровень межпопуляционной изменчивости Fst между шведскими и финскими областями подтверждает это заключение.
Из всех шведов ближайшими к финнам (диаграмма B, MDS) оказываются жители регионов Norrland и Svealand, а наиболее близкими к шведам — преимущественно шведо-говорящие жители финской области Ostrobothnia (SSOB). Что интересно, граничащие друг с другом на севере шведская область Norrbotten (NBO) и финская Northern Ostrobothnia (NOB), совершенно не показали близости на диаграмме многомерного шкалирования.
Такое положение дел может показаться странным, однако, оно полностью укладывается в исторический контекст: современное население финской области Northern Ostrobothnia (NOB) в значительной степени восходит к переселенцам из региона Southern Savonia (юго-запад Финляндии), пришедшим сюда в 16 веке. Появление мигрантов с востока страны, по видимому, нарушило более ранний градиент, существовавший в прибрежных регионах Северной Швеции и Западной Финляндии, а последующее взаимодействие шведского и финского населения было слишком слабым, чтобы этот градиент восстановить.
Для полноты картины можно рассмотреть оставшиеся пересечения между шведскими и финскими популяциями. Так, видно, к кластеру западных финнов тяготеют отдельные образцы областей NBO, VNO, ORE, STO и SMA. Первая географически находится на самом севере, вторая — на южном побережье региона Norrland, прямо напротив финской SSOB, другие три — в наиболее плотно населённой части региона Svealand, объединённой единой системой реки, в устье крупнейшей из которых находится Стокгольм.
Такая структура во всех случаях, кроме области NBO, позволяет говорить больше о шведском влиянии на финский генофонд, чем об обратном. Незначительное финское влияние можно предполагать лишь в шведской области Norrbotten (NBO). Характер влияния можно уточнить на трёхмерной диаграмме MDS (Figure S1 в статье), где первая и вторая шкала с диаграммы B дополнены третьей. Ниже приведены две проекции: шкалы 2-1 и 3-1, соответственно.

Из диаграммы видно, что образцы из Norrland (зелёные), тяготеющие к финнам по второй шкале, по третьей шкале практически полностью лежат в направлении восточных финнов. Как следствие, своим незначительным финским компонентом население шведского региона Norrland обязано по большей части финнам из соседней провинции Northern Ostrobothnia (NOB). Этот компонент появился на севере Швеции не ранее 16 века и концентрируется преимущественно в области Norrbotten (NBO).
Таким образом, один из трёх компонентов региона Norrland можно идентифицировать как позднее влияние со стороны восточных финнов, по-прежнему констатируя в целом незначительность финского влияния на население Швеции.
3. Швеция и соседние народы
Наиболее интересным оказывается сравнение шведов с соседними народами, которое авторы сопровождают несколькими любопытными диаграммами. Заключение по сравнению таково:
Общие уровни различия частоты аллелей среди населения Северной Европы выявили соответствие между генетическими и географическими расстояниями: восточные финны отличаются от остальных народов наиболее сильно; шведы — особенно из регионов Svealand и Götaland — находят наибольшее сходство с населением Центральной Европы (британцы и северные немцы). Совсем иной результат дало сравнение с образцами северных русских: наиболее далеко от них оказались шведы из региона Norrland и восточные финны, в то время как шведы из Svealand и Götaland занимают промежуточное положение, а ближайшими оказываются северные немцы и западные финны.
Это заключение подтверждают результаты многомерного шкалирования на диаграмме выше: A — сравнение народов севера Европы, B — сравнение шведов и финнов.
Из диаграммы B видно, что первая шкала ясно отделяет шведов от восточных финнов, а западные финны занимают позицию посередине. Наиболее близкими к шведам оказываются финны из прибрежных областей Ботнического залива. В то же время, сохраняется тенденция объединения в отдельный кластер шведов из региона Norrland. Не смотря на то, что образцы из шведской области Norrbotten (NBO) тяготеют в направлении финнов, дифференциация внутри шведского населения отчётливо выделяется второй шкалой: Norrland с одной стороны и Svealand/Götaland — с другой.
Что касается общей для северных европейцев диаграммы A, то — благодаря финскому участию — в её структуре нет заметных отличий от рассмотренной диаграммы B. Северные немцы из Киля занимают место в одном плотном кластере с южными шведами регионов Svealand и Götaland, шведы из Norrland по-прежнему обособлены. Однако, северные русские из Вологды оказываются между немцами с одной стороны и западными финнами с другой, будучи значительно отдалёнными от восточных финнов, на что авторы обращают особое внимание. К последнему факту они добавляют важный штрих:
В более раннем исследовании [4] мы указывали, что популяции севера Европы несут в себе различные доли структурного сходства (IBS) с населением Восточной Азии, — среди близких к ним были финны, в особенности — восточные финны. Среди шведов мы обнаружили ту же — хоть и менее выраженную — закономерность, когда образцы из региона Norrland показали наибольшую близость к Восточной Азии, а из региона Götaland — наименьшую. Такое положение дел соответствует выявленному более ранними исследованиями Y-ДНК. Существенное различие заключается в том, что ни шведы из Norrland, ни восточные финны, не показывают на малейшей близости с русскими из Вологды; вместе с тем, малое сходство между финнами и русскими можно проследить на отдельных выборках [6], [13]. Таким образом, если указанные данные являются репрезентативными для северных русских, озвученная близость жителей севера Скандинавии с населением Восточной Азии не является следствием контактов с русскими, а относится к более раннему восточному влиянию, которое предшествовало приходу славянского населения в северные регионы Восточной Европы в конце первого тысячелетия [23]. До конца не ясно, относится ли восточный компонент на территории Швеции к той же эпохе, или он явился следствием более поздних контактов с финнами.
Относительно указания авторов на наличие восточно-азиатского компонента среди восточных финнов и шведов из Norrland, следует иметь в виду выявленное выше влияние на шведов из области Norrbotten (NBO) восточно-финского компонента.
В контексте сказанного окончательно проясняется значение первой шкалы на приведённых выше диаграммах MDS (A и B), которая отчётливо разделяла шведов, немцев и русских из Вологды от западных и восточных финнов. Процесс формирования такого разительного отличия восточных финнов от своих соседей остаётся неясным. По видимому, доминирующий у восточных финнов генетический компонент был принесён отдельно и в более позднее время, в 16 веке он достиг северных областей Финляндии, а позднее появился в северной части шведского региона Norrland.
Другой интересной деталью, является внутренняя структура кластера, который на диаграмме A выше составлен шведскими и немецкими образцами. Для наглядности лучше воспользоваться трёхмерной диаграммой (Figure S1 в статье), где первая и вторая шкала с диаграммы A дополнены третьей. Ниже приведены две проекции: шкалы 2-1 и 3-1, соответственно.

Как уже указывалось, шведский регион Norrland (зелёный) ясно отделяется второй шкалой как от южной части Швеции (красный и синий), так и от немцев из Киля (оранжевый). Новыми являются детали, которые открывает третья шкала диаграммы: шведские регионы Svealand и Götaland по-прежнему находятся в едином кластере, однако немцы от них чётко отделены и находятся в противоположной части третьей шкалы — вместе с русскими из Вологды и шведами из Norrland. Не будь у образцов из Norrland такой заметной отдалённости по второй шкале, то по первой и третьей они бы оказались в одном кластере с северными немцами и северными русскими.
Другими словами, вторая и третья шкалы на диаграмме MDS описывают внутреннюю структуру народов северных регионов Европы (исключая финнов). Вторая шкала выделяет из их числа население шведского региона Norrland, а третья отделяет южных шведов от северных немцев и русских из Вологды. Любопытно здесь то, что северные шведы, северные немцы и русские оказываются в одном кластере по шкалам 1 и 3, выделяя северных шведов только по второй шкале. Остаётся сожалеть, что финские учёные скудно представили русских одними образцами из Вологды. Более масштабный анализ позволил бы ответить на большее количество вопросов, в том числе об источнике происхождения популяции региона Norrland.
4. Структурный анализ северных народов Европы
Интересностью на сладкое является диаграмма структурного анализа всех использованных SNP. Она является результатом машинного анализа и распределения на заданное количество кластеров фрагментов всех использованных в исследовании образцов. Полученные кластеры окрашиваются в различные цвета, которые соответствующим образом выделяют схожие отрезки отдельных образцов.
Ниже находится диаграмма с делением на 2...6 кластеров образцов из Британии, Киля, трёх регионов Швеции, Финляндии, Вологды и загадочного CEU, сформированного из образцов жителей североамериканского штата Юта, выходцев из Северной Европы. Образцы SWE — выборка из более раннего исследования авторов, происходящая по большей части из шведского Norrland.

Из деления на 2 кластера видно, что на диаграмме выделяется преимущественно европейские и финские компоненты. По сути, здесь налицо прямое соответствие с первой шкалой диаграмм MDS.
Деление на 3 кластера оставляет финский компонент почти неизменным, но строго разделяет остальные образцы: шведов с русскими — в одну сторону (красный), немцев с британцами — в другую (фиолетовый). Шведы из Norrland показывают наибольшую концентрацию первого компонента. Любопытно здесь и то, что бóльшую примесь первого показывают немцы, меньшую — британцы, а вторая является более характерной для южных областей Швеции, чем для северных.
В делении на 4 кластера проявляется картина, которую мы видели на диаграммах многомерного шкалирования, а именно — обособление популяций из Norrland (зелёный) по второй шкале в отдельный кластер от русских, немцев и южных шведов; незначительная финская примесь у северных шведов; заметное влияние шведов на генофонд западных финнов. Значительная концентрация зелёного компонента у северных шведов показывает, что особенность их популяции в меньшей мере является результатом смешения разнородных компонентов, и, по видимому, восходит к древнему изолированному североевропейскому населению.
Деление на 5 кластеров практически не отличается от предыдущего и дополняет картину одной деталью: появлением особого компонента, доминирующего у русских из Вологды и составляющего около половины у немцев из Киля. Незначительная примесь этого компонента наблюдается у британцев, но там она появляется лишь эпизодически, хоть в отдельных образцах и оказывается ощутимой. Вместе с тем, шведы по-прежнему представлены доминирующим красным цветом. Здесь будет уместно вспомнить третью шкалу из диаграмм MDS, которая объединяла русские и немецкие образцы воедино.
Дополнительно приводится структурная диаграмма только шведского населения, где образцы слева направо упорядочены по оси юг-север.

Резюме
1. Население Швеции показало заметную однородность. Вместе с тем, в его составе ясно выделяются два компонента, первый из которых характерен для южных регионов Götaland и Svealand, второй — для северного региона Norrland.
2. Происхождение компонента, доминирующего среди южных шведов, связано с регионами Северной Германии. Происхождение северного компонента требует уточнения, но, исходя из имеющихся данных, есть основания предполагать его длительное изолированное существование на севере Швеции и искать корни в древнейших миграциях из континентальных регионов севера Европы.
3. Финское влияние на территории Швеции является слабым, локализуется почти исключительно в северной части побережья Ботнического залива и связывается с относительно недавними миграциями восточно-финского населения.
http://drfaust-spb.livejournal.com/122197.html
Шведские гаплотипы гаплогруппы R1b1b2
Набор из 383 девятимаркерных гаплотипов Швеции, включавший 76 гаплотипов R1b1b2, был опубликован в работе [Karlsson et al, 2006].
Базовый гаплотип R1b1b2 в Швеции следующий:
13-24-14-11-11 -14-X-Y-Z-13-13-29
Это - классический Атлантический модальный гаплотип.
Все 76 шведских гаплотипов гаплогруппы R1b1b2 включают 7 базовых гаплотипов и содержат 187 мутаций. Без поправки на возвратные мутации это даёт ln(76/7)/0.017 = 140±55 поколений, и 187/76/0.017 = 145±18 поколений до общего предка. Опять мы видим почти одинаковые величины в пределах погрешности расчётов, что указывает на одного общего предка в выборке. Поправка на возвратные мутации увеличит возраст предка до 163 и 169 поколений, соответственно, то есть до 4075±1590 и 4225±520 лет до общего предка. Это - совершенно обычная цифра для европейских гаплотипов гаплогруппы R1b1b2.
Дерево гаплотипов (рис. 22.16.1) также показывает, что общий предок был один. Никакого расщепления дерева на несвязанные между собой ветви не отмечается. Некоторое сомнение может вызвать ветвь вверху справа, но там 11 гаплотипов, имеющих всего 16 мутаций, что даёт всего 91 поколение, или 2350±630 лет до общего предка. Это - просто один из более поздних вариантов генеалогической линии, в которой маркер DYS#389-1 мутировал из 13 в 14, что и сдвинуло также маркер DYS389-2, слегка изменив базовый гаплотип: 13-24-14-11-11-14-X-Y-Z-14-13-30

Рис. Дерево 9-маркерных шведских гаплотипов R1b1b2, по данным [Karlsson etal, 2006]. В выборке - 76 гаплотипов.
Это - нормальные мутации в гаплотипах, которые скорее интересны тем, кто занимается «фамильной историей».
Некоторые исландцы, возможно, являются потомками коренных жителей Америки

Бьорк — потомок эскимосов? А может, в ДНК исландцев просто сохранилась древнейшая палеоазиатская линия, равно родственная и инуитам, и лапландцам?
Первым коренным американцем, посетившим Европу, возможно, была женщина, которую захватили с собой викинги, более тысячи лет назад открывшие для себя Новый Свет.
провёл генетический анализ примерно восьмидесяти современных исландцев из четырёх семейств. Хотя большинство линий митохондриальной ДНК оказались аналогичны тем, которые наблюдаются у населения Британских островов и Скандинавии, учёным удалось обнаружить линию, относящуюся к — одной из немногих, которые участвовали в заселении Америки из Восточной Азии около 14 тыс. лет назад.
Вопреки первоначальному предположению, что эта линия проникла в Исландию вместе с недавними азиатскими иммигрантами, установлено, что последняя возможная дата появления этой линии в исландской мтДНК — 1700 год. В этой связи возникает интригующая возможность связи исландских C1-линий с путешествиями викингов в Америку в X веке.
Что самое удивительное, исландская C1-линия не принадлежит ни одной из четырёх известных субклад гаплогруппы C1, принадлежащих коренным американцам (C1b, C1c и C1d) и азиатам C1a). Пока это единственный известный член новой субклады C1e.
Исследователи просят воздержаться от далекоидущих выводов о том, что исландцы произошли от американоидов, или о том, что часть исландцев является потомками американцев, возможно, вывезенных викингами. Хотя C1e, скорее всего, имеет американское происхождение, нельзя исключать азиатские и европейские корни этой линии. Например, можно предположить, что C1e просто мало распространена в Европе, а в Исландии её частотность повысилась в условиях изоляции сравнительно небольшого населения.

Результаты исследования опубликованы в издании .
Подготовлено по материалам
.
|












![[pc1pc2.png]](https://blogger.googleusercontent.com/img/b/R29vZ2xl/AVvXsEisNYsSED1P18IIetjiglDBxAp8NXSq9F2boS2n3T68yhFeNkTyWTduvdjZL_AOokGDTXy2i_1bQTKy3UabVQmwjds6OTBsvpq4iGhcv7b5TAp1CUryYygXGy3V37IK-fgOlbHw0Lw__F8/s1600/pc1pc2.png)



