ФЕДОРОВА Сардана Аркадьевна
ОБЩАЯ ХАРАКТЕРИСТИКА РАБОТЫ
Актуальность темы. Исследования разнообразия генома человека в популяциях получили бурное развитие за последние годы в связи с полной расшифровкой генома в начале XXI в. Первоначально анализ особенностей геномного полиморфизма был направлен на решение вопросов происхождения современного человека и эволюции генома в целом. Стремительный прогресс в этой области знаний позволил определить основные пути заселения человеком территории земного шара (Cann et al., 1987; Vigilant et al., 1991; Ingman et al., 2000; Kivisild et al., 2003; Macaulay et al., 2005; Metspalu et al., 2006; Seielstad et al., 1999; Hammer et al., 1998, 2001; Underhill et al., 2000, 2001a), и в настоящее время особую актуальность приобретают работы, связанные с более глубоким изучением эволюционной и демографической истории отдельных регионов. Значительное число исследований проведено к настоящему времени по популяциям Западной и Восточной Европы (Torroni et al., 1994d, 1996, 1998; Richards et al., 1998, 2002; Macaulay et al., 1999b; Finnila et al., 2001; Herrnstadt et al., 2002; Tambets, 2004; Loogvali et al., 2004; Semino et al., 2000, 2004; Rosser et al., 2000; Zerjal et al., 2001; Cavalli-Cforza and Minch, 1997; Лимборская, 2002; Хуснутдинова, 1999, 2006; Бермишева, 2002; Malyarchuk and Derenko, 2001; Belyaeva et al., 2003; Malyarchuk et al., 2004; Balanovsky et al., 2008), Ближнего Востока (Richards et al., 2000, 2002, 2003; Luis et al., 2004; Semino et al., 2004; Al-Zahery et al., 2003; Shlush et al., 2008), Индии (Passarino et al., 1996; Kivisild et al., 1999, 2003; Bamshad et al., 2001; Palanichamy et al., 2004; Sengupta et al., 2006), Восточной Азии (Su et al., 1999; Jin and Su, 2000; Karafet et al., 2001; Kivisild et al., 2002; Kong et al., 2003a, 2003b, 2006; Yao et al., 2002a, 2002b, 2002c, 2004; Tanaka et al., 2004), Сибири и Средней Азии (Karafet et al., 2002; Derenko et al., 2003, 2006, 2007; Степанов, 2002, 2006; Харьков, 2005; Starikovskaya et al., 1998, 2005; Schurr et al., 1999; Derbeneva et al., 2002; Дербенева, 2002; Volodko et al., 2008; Comas et al., 1998, 2004; Wells et al., 2001; Quintana-Murci et al., 2004), Америки (Torroni et al., 1992, 1993a, 1993b, 1994a, 1994c; Karafet et al., 1999; Zegura et al., 2004; Карафет, 2006; Tamm et al., 2007). Относительно небольшое количество работ посвящено популяциям Якутии. Лишь в последние годы появились публикации, касающиеся генетической истории якутов (саха) (Pakendorf et al., 2002, 2003, 2006; Федорова, 2003; Пузырев, 2003; Хитринская, 2003; Tarskaya et al., 2006; Тарская и Мелтон, 2006; Харьков, 2008; Zlojutro et al., 2008) и юкагиров (Volodko et al., 2008). Масштабные этногеномные исследования, в которых генофонд коренных народов Якутии рассматривался бы как целостная система, до сих пор не проводились.
Республика Саха (Якутия), расположенная на северо-востоке Российской Федерации, является одним из интереснейших регионов мира по своеобразию процессов формирования генофонда коренного населения. Территория республики составляет 18% от территории Российской Федерации, но в силу суровых природных и климатических условий слабо заселена. На огромном пространстве, 4/5 которого занято тайгой, с узкой полосой тундры на севере, проживает менее 1 млн. человек. Коренное население Якутии (якуты, эвенки, эвены, юкагиры, долганы, чукчи), по материалам последней Всероссийской переписи 2002 г., составляет 49% от общей численности. Особенностями популяций Якутии являются немногочисленность и рассеянность на огромной территории. Коренное население Якутии неоднородно по антропологической и языковой принадлежности, культуре, традициям, типам хозяйства.
Археологические данные указывают на то, что территория Якутии была заселена с древнейших времен. Наиболее древние стоянки человека эпохи верхнего палеолита были открыты на реках Алдан (35-33 тыс. лет назад) (Мочанов, 1977) и Яна (30 тыс. лет назад) (Pitulko et al., 2004). Близкое типологическое сходство изделий дюктайской эпохи (35-10,5 тыс. лет назад) с древнейшими палеоиндейскими позволило выдвинуть гипотезу о генетической связи дюктайцев с предками американских индейцев, проникших на американский континент сухопутным путем через Чукотку и Берингию (Мочанов, 1977). Вопрос этнической идентификации древнего населения Якутии новокаменного века до сих пор остается наиболее спорным моментом в исторических реконструкциях: с древними неолитическими племенами Якутии связывают все известные современные субарктические этносы - юкагиров, эвенов, нганасанов, чукчей, эскимосов и коряков (Окладников, 1955a, 1970; Федосеева, 1980; Константинов, 1978; Алексеев, 1996a, 1996б). Позднее, начиная с железного века, территория Якутии, по-видимому, стала заселяться предками современных эвенков, вначале пешими и затем оленеводческими группами (Константинов, 1978; Туголуков, 1980, 1985). В результате взаимодействия предков эвенков с древними юкагирами и коряками возник новый тунгусоязычный этнос - эвены. Тюркоязычные скотоводческие племена предков современных якутов (саха) рассматриваются как наиболее поздние мигранты, переселившиеся в бассейн средней Лены из Прибайкалья. По вопросу о времени миграции якутов на север во мнениях историков имеются значительные расхождения: с I по XII вв. (Ксенофонтов, (1937), 1992), с VI в. н.э. (Алексеев, 1996б), c X по XVI вв. (Окладников, 1955а), не ранее XIII-XIV вв. (Гоголев, 1993), с XV в (Константинов, 1975). Самый молодой этнос Сибири - долганы сложился лишь к XIX в., вобрав линии якутов, эвенков, русских, энцев и ненцев (Долгих, 1963).
Первые этногеномные исследования, в которых затрагивались отдельные популяции Якутии, были сфокусированы на изучении древних миграций человека по территории Евразии (Zerjal et al., 1997; Torroni et al., 1998; Lahermo et al., 1999) и заселения Америки (Torroni et al., 1993a; Karafet et al., 1999). Более интенсивные исследования были предприняты по изучению структуры генофонда якутов как наиболее многочисленного этноса Сибири, однако до сих пор не удалось получить однозначной генетической оценки по вопросам происхождения и генетической истории народа саха. Расхождения во мнениях авторов касаются в основном соотношения «пришлых» и автохтонных элементов в генофонде якутов и степени генетического родства с другими этносами Сибири. Противоречия определяются, на наш взгляд, главным образом, недостаточной изученностью структуры генофонда непосредственных соседей якутов - эвенков, эвенов и юкагиров, параметров внутриэтнической вариабельности, а также недостаточной глубиной филогенетического анализа для всех изученных к настоящему времени популяций.
В представляемой работе для анализа разнообразия генома в популяциях мы использовали несколько молекулярно-генетических систем - митохондриальную ДНК (мтДНК), Y-хромосому и аутосомные Alu-инсерции. Исключительная информативность первых двух систем обусловлена такими свойствами, как специфичный характер наследования - по материнской и отцовской линиям, отсутствие рекомбинационных процессов и высокая степень полиморфности. Реконструкция филогенетических деревьев мтДНК и Y-хромосомы с учетом времени коалесценции и в совокупности с географическими и археологическими данными составляет основу филогеографического подхода, который является наиболее перспективным в популяционных исследованиях в настоящее время (Avise, 2000). В отличие от локусов ядерной ДНК, где эволюционные изменения прослеживаются главным образом по вариабельности частот различных аллелей в популяциях, митохондриальная ДНК и Y-хромосома дают возможность восстановить действительную филогению, т.е. последовательность возникновения носителей различных гаплотипов в эволюционном ряду. Анализ структуры и распределения линий в различных регионах позволяет определить направленность миграционных потоков, имевших место в прошлом. Для более глубокого понимания происхождения и формирования генофонда отдельных этносов и процессов эволюции генома человека в целом необходимы детальные филогеографические исследования концевых ветвей филогенетических деревьев митохондриальной ДНК и Y хромосомы, что становится возможным с использованием большего количества информативных ДНК-маркеров, с развитием новых подходов к анализу данных, и накоплением фактологического материала.
Значительный интерес представляет сравнительный анализ линий в древних и современных популяциях. Молекулярно-генетический анализ костных останков из древних погребений базируется главным образом на исследовании митохондриальной ДНК, что обусловлено ее многокопийностью и, как следствие, значительно большей сохранностью в скелетном материале по сравнению с ядерной ДНК. В большинстве опубликованных работ результаты исследования ДНК популяций неолита и палеолита свидетельствуют о генетической преемственности между древним и современным населением (Oota et al., 1998, 2001b; Gonzales-Oliver et al., 2001; Jones, 2003; Garcia-Bour et al., 2004; Vernesi et al., 2004; Adachi et al., 2004; Ricaut et al., 2004; Keyser-Tracqui et al., 2006). Исследования ДНК человека из древних погребений Якутии немногочисленны - известны единичные публикации французских исследователей, где представлен анализ мтДНК неолитической шаманки (Ricaut et al., 2005), мужчины из погребения раннего железного века (Amory et al., 2006), 8 останков из якутских погребений XVII и XVIII вв. (Ricaut et al., 2004, 2006).
Изучение геномного разнообразия имеет значение не только для решения вопросов происхождения и генетической истории различных этносов, но также является основой для молекулярной эпидемиологии наследственных и мультифакториальных заболеваний. Каждый регион характеризуется определенным набором наиболее распространенных болезней. Для понимания причин распространенности тех или иных заболеваний в различных регионах, а также для разработки подходов к их ранней ДНК-диагностике и эффективной профилактике первоначально необходимо проведение популяционных исследований по ДНК-локусам, определяющим развитие заболевания. Такие разработки особенно актуальны и перспективны в Якутии в связи с особенностями распространения некоторых наследственно-обусловленных болезней в регионе (Ноговицына, 1999; Назаренко, 2002а, 2002б; Платонов, 2003; Тарская, 2004; Тарская и Ельчинова, 2006; Сухомясова, 2005; Галеева, 2006; Максимова, 2007; Maksimova et al., 2007) .
В связи с вышеизложенным были определены цель и задачи данного исследования.
Цель исследования: охарактеризовать структуру генофонда и генетические взаимоотношения популяций Якутии по данным полиморфизма митохондриальной ДНК, Y-хромосомы и аутосомных ДНК-локусов.
Задачи исследования:
1. определить особенности структуры генофонда коренного населения РС(Я) с помощью трех молекулярно-генетических систем - митохондриальной ДНК, Y-хромосомы и аутосомных Alu-инсерций, провести сравнительный анализ с популяциями соседних регионов;
2. определить соотношение западно- и восточноевразийских линий мтДНК и Y-хромосомы в генофонде народов РС(Я);
3. оценить параметры генетического разнообразия и степень генетической дифференциации популяций Якутии по данным полиморфизма мтДНК, Y-хромосомы и аутосомных Alu-инсерций; 4. провести филогеографический анализ наиболее распространенных в популяциях РС(Я) гаплогрупп мтДНК и Y хромосомы;
5. охарактеризовать генетические взаимоотношения между изученными популяциями и определить положение генофонда населения Якутии в системе генофондов популяций соседних регионов по данным полиморфизма мтДНК и Y-хромосомы;
6. провести анализ мтДНК из древних погребений Якутии;
7. провести анализ полиморфизма локусов, сцепленных с генами
наследственных болезней, в популяциях Якутии.
Научная новизна. Впервые проведено исследование структуры генофонда народов Якутии (якуты, эвенки, эвены, юкагиры и долганы) как целостной популяционной системы с использованием оценки генетического разнообразия митохондриальной ДНК, Y-хромосомы, аутосомных Alu-инсерций и высокополиморфного участка (CTG)n-повторов DMPK-гена. На основании данных об изменчивости линий митохондриальной ДНК и Y хромосомы в популяциях Якутии получены детальные характеристики о структуре генофонда коренного населения РС(Я). Определено соотношение западно- и восточноевразийских линий мтДНК и Y хромосомы в генофондах народов Якутии, проведены оценки уровня генетического разнообразия и степени генетической дифференциации популяций региона в целом. Проведен филогеографический анализ мажорных гаплогрупп С и D5а2 митохондриальной ДНК и гаплогруппы N Y хромосомы. Получены новые данные по разнообразию типов митохондриальной ДНК из древних погребений Якутии эпохи средневековья и позднего неолита. Установлено положение генофонда народов Якутии в системе генофондов популяций соседних регионов (Чукотки, Камчатки, Дальнего Востока, Северного Китая, Монголии, Южной и Западной Сибири). Полученные в работе данные существенно дополняют и расширяют представления о путях миграций популяций человека и общей картине заселения Северо-Восточной Евразии.
Научно-практическая значимость. Результаты исследования генетической истории коренных этносов Якутии представляют значительный интерес для специалистов смежных отраслей знания - историков, этнографов, археологов, антропологов и лингвистов и могут использоваться при решении проблем этногенеза коренных народов Якутии. Полученные данные имеют важное значение для понимания причин накопления некоторых наследственных заболеваний в регионе. Данные по генетическому полиморфизму могут иметь существенное значение в области судебной медицины, геногеографии, эпидемиологии наследственных и мультифакториальных заболеваний. Созданные в целях выполнения работы коллекции ДНК популяций РС(Я) могут использоваться в дальнейшем для проведения популяционных, эволюционных и медико-генетических исследований. Материалы работы могут быть использованы в научно-образовательном процессе при создании курсов лекций для студентов биологических, медицинских, исторических специальностей, а также для издания научно-популярной литературы и в средствах массовой информации.
Основные положения, выносимые на защиту.
1. Спектр и частоты гаплогрупп мтДНК и Y хромосомы, аллельных вариантов (CTG)n-локуса гена DMPK, частоты 8 аутосомных Alu-инсерций в популяциях Якутии. Характерные особенности структуры генофонда якутов, эвенков, эвенов, юкагиров и долган.
2. Выраженность восточноевразийского компонента, низкое содержание западноевразийских линий мтДНК и Y-хромосомы в генофонде популяций Якутии. Присутствие палеоевропеоидного компонента в генофонде якутов и эвенков;
3. Тесное генетическое родство между центральными и вилюйскими якутами, большая генетическая близость популяций якутов к эвенкам Якутии и отдаленность якутов от юкагиров по данным полиморфизма мтДНК и Y-хромосомы;
4. Большая выраженность отличий генетических портретов популяций Якутии по линиям Y хромосомы, чем по линиям мтДНК, что определяется в основном эффектом основателя в популяциях якутов и эффектом патрилокальности; незначительная степень дифференциации между популяциями якутов и эвенков Якутии по полиморфизму мт-ДНК и Alu-инсерционных локусов;
5. Заселение территории Якутии из регионов Южной Сибири, Монголии и Северного Китая; относительная изолированность от восточных регионов Чукотки и Камчатки по данным полиморфизма мтДНК, начиная по крайней мере с эпохи позднего неолита;
6. Преемственность линий мтДНК в популяции якутов за последние 300 лет;
7. Эффект основателя или наличие в генах, ответственных за наследственные болезни, одной мажорной мутации для наследственных болезней, имеющих редкую частоту возникновения и получивших распространение в популяции якутов.
Апробация работы. Материалы исследования были представлены в виде докладов (устных и стендовых) или тезисами докладов на II и III Международной конференциях «Проблемы вилюйского энцефаломиелита и других нейродегенеративных заболеваний в Якутии» (Якутск, 2000, 2006), 7th Annual Human Genom Meeting (Shanghai, 2002), республиканских научно-практических конференциях «Актуальные вопросы детской неврологии и педиатрии» (Якутск, 2001), «Достижения и перспективы медицинской науки в Республике Саха (Якутия)» (Якутск, 2001), «Вопросы формирования здоровья и патологии человека на Севере: факты, проблемы и перспективы» (Якутск, 2002), на Всероссийском научно-практическом симпозиуме «Технологии генодиагностики в практическом здравоохранении» в рамках Международной конференции «Геномика, протеомика и биоинформатика для медицины» (Москва, 2002), 13th ingress of the European anthropological association (Zagreb, Croatia, 2002), Meeting Human Origin and Diseases (Cold Spring Harbor, 2002), 2d International Meeting of genetic of complex diseases and isolated populations (Italy, 2003), Всероссийской научно-практической конференции «Современные достижения клинической генетики» (Москва, 2003), Х Российско-Японском Международном симпозиуме медицинского обмена (Якутск, 2003), 9th Annual Human Genom Meeting (Berlin, 2004), V Оьезде Российского общества медицинских генетиков (Уфа, 2005), 10th Annual Human Genom Meeting (Kioto, 2005), Международной конференции «Генетические аспекты патологии человека. Проблемы сохранения генофонда коренных народов Севера» (Якутск, 2005), 55th Annual Meeting of American Society of Human Genetics (Uta, 2005), межрегиональной научно-практической конференции «Молекулярно-клеточные аспекты патологии человека на Севере» (Якутск, 2007), форуме молодых ученых РС(Я) (Якутск, 2008), научных семинарах ЯНЦ СО РАМН и Эстонского Биоцентра, семинаре «Популяционная и эволюционная генетика» Отдела генетики популяций и природопользования ИОГен РАН (апробация диссертации, 23 мая 2007 г.).
Декларация личного участия автора. В диссертации использованы экспериментальные данные, полученные лично автором: гаплотипирование мтДНК и Y хромосомы осуществлено на базе Эстонского Биоцентра (г.Тарту, Эстония), анализ мтДНК из костных останков проведен в лаб. геобиологии Университета Нагойя (г. Нагойя, Япония) и в Эстонском Биоцентре, анализ полиморфизма (CTG)n-локуса DMPK-гена в популяциях РС(Я) и скрининг мутаций в гене гемохроматоза в популяции якутов выполнен в Отделе молекулярной генетики ЯНЦ СО РАМН. Автор лично участвовал в проведении трех экспедиций в северные улусы, организации Банка ДНК ЯНЦ СО РАМН, внедрении в практику медико-генетического консультирования методов ДНК-диагностики 6 наследственных болезней, распространенных в РС(Я). Генотипирование Alu-инсерций проведено сотрудниками Института биохимии и генетики УНЦ РАН (г. Уфа, Россия). Работа выполнена в рамках реализации Республиканской целевой программы «Развитие генодиагностики человека в РС(Я)» согласно Договорам о научно-техническом сотрудничестве между ЯНЦ СО РАМН, ИБГ УНЦ РАН, Эстонским Биоцентром и лаб. геобиологии Университета Нагойи. Суммарно личное участие автора составило ~75%.
Благодарности. Автор бесконечно признателен людям, предоставившим возможность для выполнения данного исследования - своему научному консультанту, члену-корреспонденту АН Республики Башкортостан, заведующей отделом геномики Института биохимии и генетики УНЦ РАН профессору Эльзе Камилевне Хуснутдиновой, президенту АН Эстонии, директору Эстонского Биоцентра профессору Рихарду Виллемсу, директору ЯНЦ СО РАМН профессору Михаилу Иннокентьевичу Томскому, заместителю директора по научной работе
ЯНЦ СО РАМН (2002-2007) профессору Валерию Архиповичу Аргунову, директору Департамента по охране генофонда народов РС(Я) (1994-2002) Валентине Ивановне Кириллиной, заведующему лабораторией геобиологии Университета Нагойя профессору Tomowo Ozawa, заместителю директора ФГНЦ «Институт здоровья» РС(Я) профессору Федору Алексеевичу Платонову.
Автор выражает искреннюю благодарность сотрудникам Эстонского Биоцентра Маре Рейдла, Юрию Парик, Эне Метспалу, Сиири Рутси, Марии Адоаян, сотрудникам Института биохимии и генетики УНЦ РАН Ильдусу Альбертовичу Кутуеву, Рите Игоревне Хусаиновой, Вите Леоновне Ахметовой, Марине Александровне Бермишевой, Баязиту Булатовичу Юнусбаеву за методическую помощь и ценные советы при проведении данного исследования, заместителю директора Музея археологии ЯГУ Александру Дмитриевичу Степанову за предоставление костного материала поздненеолитического Кёрдюгенского погребения, ценные замечания и обсуждение результатов работы, археологу Эдуарду Константиновичу Жиркову за предоставление костного материала из якутских погребений.
Автор считает приятным долгом поблагодарить зав. отделом молекулярной генетики (2002-2006) Анну Николаевну Ноговицыну за организационную помощь. Автор выражает сердечную благодарность сотрудникам отдела молекулярной генетики ЯНЦ СО РАМН и врачам Медико-генетической консультации РБ№1-НЦМ, принимавшим участие в экспедициях, а также всем донорам, давшим согласие на предоставление материала. Отдельную благодарность хотелось бы выразить сотрудникам лаборатории молекулярной генетики, людям, оказавшим дружескую поддержку в течение всего времени выполнения работы.
Публикации. В диссертации обобщены данные 50 работ, включая 1 монографию, 15 статей в отечественных и зарубежных журналах, 34 тезисов в материалах конференций.
Структура и объем работы. Диссертация изложена на 338 страницах машинописного текста, состоит из введения, пяти глав обзора литературы, описания материалов и методов исследования, семи экспериментальных глав, заключения, выводов и библиографического списка использованной литературы. Работа иллюстрирована 35 таблицами, 37 рисунками и дополнена 7 приложениями. Список литературы включает 507 публикаций отечественных и зарубежных авторов.
МАТЕРИАЛЫ И МЕТОДЫ
Коллекции ДНК для популяционных исследований составлены из материала банка ДНК отдела молекулярной генетики ЯНЦ СО РАМН. В популяционные выборки включались здоровые неродственные индивиды, этническая принадлежность которых учитывалась преимущественно до третьего поколения. Лингвистические и антропологические характеристики исследованных этнических групп, а также численность изученных выборок по мтДНК, Y хромосоме, участку (CTG)n-повторов гена DMPK приведены в табл.1.

Анализ Alu-инсерций был проведен в смешанной выборке центральных и вилюйских якутов (n=85), и в популяции эвенков Жиганского улуса (n=41). При скрининге мутаций Cys282Tyr и His63Asp гена гемохроматоза (HFE) была использована смешанная выборка центральных и вилюйских якутов (n=84).
Древняя ДНК выделена из костных останков могильников центральной Якутии: трех человек из якутских погребений (Ат-Дабаан, Одьулуун, Дьараама) (XVIII в.) и двух человек из поздненеолитического Кёрдюгенского погребения тыс. до н.э.). В работе были использованы следующие костные останки: образец №1 выделен из коренного зуба (М ), №2 - из плечевой кости (Trochlea humeri), №3 -костей предплечья (Radius, Ulna), №4 - коленной чашечки (Patella), №5 -ключичной кости (Qavicula). Для исследования брали внутреннее содержимое костей (3-5 г), которое разрушали до порошкообразного состояния в металлических ступках, предварительно стерилизованных прокаливанием при 2000C в течение 2 ч. ДНК экстрагировали из костного порошка с использованием набора GENECLEAN Kit for Ancient DNA (BIO 101, USA). Гаплотипирование линий мтДНК осуществляли путем секвенирования ГВО(16024-16390) мтДНК, ПДРФ-анализа и секвенирования 40 диагностических сайтов кодирующей области. Фрагменты контрольного региона мтДНК, содержащие ГВСI и ГВСII, амплифицировали и очищали с применением набора ExoSAP-IT "Applied Biosystems". Амплификацию проводили с использованием праймеров, описанных ранее (Cavellier et al., 2000). Нуклеотидные последовательности ГВС (16024—16390 п.н.) и ГВСП (70-350) мтДНК секвенировали с помощью DYEnamictm ET Terminator Cycle Sequencing Kit ("Amersham Pharmacia Biotech", Швеция) и анализировали на автоматических секвенаторах Applied Biosystems 377 DNA Sequencer и MegaBACE1000 Sequencer.
Выравнивание и анализ нуклеотидных последовательностей проводили с использованием пакета прикладных программ Seqlab (Genetics Computer Group Wisconsin Package Version 10.0) и ChromasPro, version 1.41. Последовательности ГВСП между позициями 70-350 определяли только для некоторых линий гаплогрупп F2a, M7, M13 и R*. Нуклеотидные замены в позициях 3206, 3816, 7933, 8020, 9181, 10646, 11078, 11944, 12026, 12672, 12705, 12930, 13104 и 15204 ГВа анализировали путем секвенирования. Для полиморфных участков 633, 3010, 4833, 4958, 5178, 5301, 7028, 7196, 7476, 7853, 7933, 8414, 8994, 9055, 9824, 10034, 10398, 10400, 12308, 12406, 12950, 13101, 13263, 13368, 13708 и 14766 проводили ПДРФ- анализ c помощью соответствующих рестриктаз.
Принадлежность к определенным гаплогруппам устанавливали, сравнивая анализируемую нуклеотидную последовательность ГВСI с кембриджской эталонной последовательностью (Andrews et al., 1999) и классифицировали согласно современной номенклатуре западно- и восточноевразийских гаплогрупп мтДНК (Torroni et al., 1992, 1994, 1996; Richards et al., 1998; Macaulay et al., 1999; Schurr et al., 1999; Finnila et al., 2001; Herrnstadt et al., 2002; Kivisild et al., 2002; Yao et al., 2004; Kong et al, 2003; Comas et al., 2004; Tanaka et al., 2004; Kong et al. 2006; Derenko et al., 2007).
Филогенетические сети конструировали в программе Network 4.2.0.1 (http://www.fluxus-engineering.com) с последовательным использованием алгоритмов reduced median (Bandelt et al., 1995), и median joining (Bandelt et al., 1999). Позициям в ГВСI присваивался различный «вес» (1-4) в зависимости от частоты возникновения мутаций в процессе эволюции (Allard et al., 2002). Время коалесценции (р) и доверительный интервал вычисляли согласно общепринятым формулам (Forster et al., 1996; Saillard et al., 2000).
Гаплотипы Y хромосомы идентифицировали путем анализа 24 диалелльных маркеров нерекомбинирующей области. Маркеры М9, TAT, SRY1532, 92R7, M20, M89, P43, P37, M231, M35, M78 тестировали методом ПДРФ-анализа согласно опубликованным протоколам (Raitio et al., 2001; Zerjal et al., 1997; Whitfield et al., 1995; Underhill et al., 1997, 2000, 2001; Mathias et al., 1994; Akey et al., 2001; Karafet et al., 2002). Полиморфные позиции в локусах M48, M52, M70, M130, M170, M173, M175, M201, M217, М253 и M269 анализировали секвенированием соответствующих фрагментов ДНК (Underhill et al., 2000, 2001а; Cruciani et al., 2002). Маркеры YAP и 12f2 анализировали согласно Hammer and Horai (1995) и Casanova et al. (1985) соответственно. Использовали номенклатуру гаплогрупп Y хромосомы, предложенную Международным консорциумом (YCC, 2002) и последний вариант номенклатуры, предложенный Т.Карафет и соавт. (2008).
6 микросателлитных локусов (DYS19, DYS388, DYS390, DYS391, DYS392, DYS393) анализировали по отдельности путем амплификации и последующего анализа ПЦР-продуктов на секвенаторе MegaBACE1000. Для определения длины фрагмента и числа повторов использовали программу Genetic Profiler 2.2. Время коалесценции N3 (N1C)- и С3с-гаплотипов по вариабельности микросателлитных маркеров определяли согласно L.Zhivotovsky et al., 2004.
При анализе древней ДНК методом гнездовой ПЦР амплифицировали три перекрывающихся фрагмента ГВа 15986-16149, 16106-16271 и 16209-16401. Продукты амплификации разделяли электрофорезом в 3%-легкоплавкой агарозе, элюировали из геля фрагменты ДНК соответствующей длины с последующей очисткой и переосаждением в этаноле. Полученные фрагменты ДНК секвенировали с обеих цепей с прямыми и обратными праймерами (Yang et al., 1999). Фрагменты ГВСТ мтДНК, результаты секвенирования которых были неоднозначны, клонировали с использованием набора InsT/Aclone PCR Product Cloning Kit, Fermentas. Для каждого фрагмента анализировали несколько бактериальных клонов, содержащих вставку соответствующей длины.
Генотипирование локусов, содержащих Alu-инсерционные полиморфизмы (ACE, APOA1, PV92, TPA25, Ya5NBC27, Ya5NBC102, Ya5NBC148, Ya5NBC182) проводили согласно опубликованным ранее протоколам (Arcot et al. 1996, 1997, 1998; Stoneking et al. 1997; Caroll et al., 2001).
Анализ числа CTG-повторов DMPK гена проводили с применением праймеров, описанных ранее (Brook et al., 1992). Генотипирование Cys282Tyr и His63Asp мутаций гена HFE проводилось по соответствующим протоколам (Beutler et al., 1996).
Статистический анализ базировался на частотах линий и гаплогрупп мтДНК и Y-хромосомы. 95%-ные доверительные интервалы для частот гаплогрупп рассчитывали в программе Sampling, любезно предоставленной V.Macaulay и адаптированной М.Мetspalu (Эстонский Биоцентр). Для оценки генетических взаимоотношений между популяциями использовали метод главных компонент, кластерный анализ и определение вероятности совпадения линий между популяциями. Для проведения факторного анализа с применением метода главных компонент использовали программу POPSTR, любезно предоставленную H.Harpending (Эстонский Биоцентр), и пакет прикладных программ Statistica, version 5.5. (StatSoft Inc., USA). Для проведения кластерного анализа были использованы матрицы генетических расстояний Fst (Reynolds et al., 1983), рассчитанные в пакете программ ARLEQUIN, version 3.01. Филогенетические деревья строили по методу связывания ближайших соседей (neighbor-joining) с использованием программы MEGA, version 2.1. Вероятность совпадения линий (m) между популяциями определяли по формуле: m=∑xiyi, где xi и yi - частоты совпадающей линии i в популяциях X и Y, соответственно.
Расчет показателей генетического разнообразия (h) (Nei, 1987) и анализ молекулярной дисперсии (AMOVA, Excoffier et al., 1992) проводили в пакете программ ARLEQUIN, version 3.01. Значимость Fst оценивали числом пермутаций 10000. Степень генетической дифференциации между изученными популяциями оценивали также с помощью стандартизированной Gst (Gst'), предложенной P.W.Hedrick (Hedrick, 2005), которая позволяет проводить сравнение между локусами с различной степенью генетической вариабельности и скоростью мутирования.
Тестирование соответствия распределения генотипов закону Харди-Вайнберга по Alu-инсерционным локусам, расчет фактической и теоретической гетерозиготности проводили в программе GenePop, version 3.3. Степень генетической дифференциации между популяциями по Alu-инсерционным локусам оценивали с помощью показателя Gst, который рассчитывали по Нею (Nei, 1983). Анализ потока генов проводили в соответствии с моделью, предложенной KHarpending and R.Ward (1982).
Статистический анализ полученных данных по микросателлитным локусам проводили с использованием компьютерных программ Microsat и GenPop (version 3.3). Для оценки генетических расстояний между популяциями использовали меру Sfi (Goldstein et al., 1995). Тестирование на соответствие распределения частот генотипов закону Харди-Вайнберга проводили по модифицированному методу цепи Маркова (Guo and Thompson, 1992).
РЕЗУЛЬТАТЫ И ОБСУЖДЕНИЕ 1. Структура и происхождение митохондриального генофонда популяций Якутии
1.1. Особенности структуры митохондриального генофонда популяций Якутии. Филогенетическая сеть гаплотипов митохондриальной ДНК в популяциях Якутии (n=694) показана на рис.1.
Древняя ДНК выделена из костных останков могильников центральной Якутии: трех человек из якутских погребений (Ат-Дабаан, Одьулуун, Дьараама) (XVIII в.) и двух человек из поздненеолитического Кёрдюгенского погребения тыс. до н.э.). В работе были использованы следующие костные останки: образец №1 выделен из коренного зуба (М ), №2 - из плечевой кости (Trochlea humeri), №3 -костей предплечья (Radius, Ulna), №4 - коленной чашечки (Patella), №5 -ключичной кости (Qavicula). Для исследования брали внутреннее содержимое костей (3-5 г), которое разрушали до порошкообразного состояния в металлических ступках, предварительно стерилизованных прокаливанием при 2000C в течение 2 ч. ДНК экстрагировали из костного порошка с использованием набора GENECLEAN Kit for Ancient DNA (BIO 101, USA). Гаплотипирование линий мтДНК осуществляли путем секвенирования ГВО(16024-16390) мтДНК, ПДРФ-анализа и секвенирования 40 диагностических сайтов кодирующей области. Фрагменты контрольного региона мтДНК, содержащие ГВСI и ГВСII, амплифицировали и очищали с применением набора ExoSAP-IT "Applied Biosystems". Амплификацию проводили с использованием праймеров, описанных ранее (Cavellier et al., 2000). Нуклеотидные последовательности ГВС (16024—16390 п.н.) и ГВСП (70-350) мтДНК секвенировали с помощью DYEnamictm ET Terminator Cycle Sequencing Kit ("Amersham Pharmacia Biotech", Швеция) и анализировали на автоматических секвенаторах Applied Biosystems 377 DNA Sequencer и MegaBACE1000 Sequencer.
Выравнивание и анализ нуклеотидных последовательностей проводили с использованием пакета прикладных программ Seqlab (Genetics Computer Group Wisconsin Package Version 10.0) и ChromasPro, version 1.41. Последовательности ГВСП между позициями 70-350 определяли только для некоторых линий гаплогрупп F2a, M7, M13 и R*. Нуклеотидные замены в позициях 3206, 3816, 7933, 8020, 9181, 10646, 11078, 11944, 12026, 12672, 12705, 12930, 13104 и 15204 ГВа анализировали путем секвенирования. Для полиморфных участков 633, 3010, 4833, 4958, 5178, 5301, 7028, 7196, 7476, 7853, 7933, 8414, 8994, 9055, 9824, 10034, 10398, 10400, 12308, 12406, 12950, 13101, 13263, 13368, 13708 и 14766 проводили ПДРФ- анализ c помощью соответствующих рестриктаз.
Принадлежность к определенным гаплогруппам устанавливали, сравнивая анализируемую нуклеотидную последовательность ГВСI с кембриджской эталонной последовательностью (Andrews et al., 1999) и классифицировали согласно современной номенклатуре западно- и восточноевразийских гаплогрупп мтДНК (Torroni et al., 1992, 1994, 1996; Richards et al., 1998; Macaulay et al., 1999; Schurr et al., 1999; Finnila et al., 2001; Herrnstadt et al., 2002; Kivisild et al., 2002; Yao et al., 2004; Kong et al, 2003; Comas et al., 2004; Tanaka et al., 2004; Kong et al. 2006; Derenko et al., 2007).
Филогенетические сети конструировали в программе Network 4.2.0.1 (http://www.fluxus-engineering.com) с последовательным использованием алгоритмов reduced median (Bandelt et al., 1995), и median joining (Bandelt et al., 1999). Позициям в ГВСI присваивался различный «вес» (1-4) в зависимости от частоты возникновения мутаций в процессе эволюции (Allard et al., 2002). Время коалесценции (р) и доверительный интервал вычисляли согласно общепринятым формулам (Forster et al., 1996; Saillard et al., 2000).
Гаплотипы Y хромосомы идентифицировали путем анализа 24 диалелльных маркеров нерекомбинирующей области. Маркеры М9, TAT, SRY1532, 92R7, M20, M89, P43, P37, M231, M35, M78 тестировали методом ПДРФ-анализа согласно опубликованным протоколам (Raitio et al., 2001; Zerjal et al., 1997; Whitfield et al., 1995; Underhill et al., 1997, 2000, 2001; Mathias et al., 1994; Akey et al., 2001; Karafet et al., 2002). Полиморфные позиции в локусах M48, M52, M70, M130, M170, M173, M175, M201, M217, М253 и M269 анализировали секвенированием соответствующих фрагментов ДНК (Underhill et al., 2000, 2001а; Cruciani et al., 2002). Маркеры YAP и 12f2 анализировали согласно Hammer and Horai (1995) и Casanova et al. (1985) соответственно. Использовали номенклатуру гаплогрупп Y хромосомы, предложенную Международным консорциумом (YCC, 2002) и последний вариант номенклатуры, предложенный Т.Карафет и соавт. (2008).
6 микросателлитных локусов (DYS19, DYS388, DYS390, DYS391, DYS392, DYS393) анализировали по отдельности путем амплификации и последующего анализа ПЦР-продуктов на секвенаторе MegaBACE1000. Для определения длины фрагмента и числа повторов использовали программу Genetic Profiler 2.2. Время коалесценции N3 (N1C)- и С3с-гаплотипов по вариабельности микросателлитных маркеров определяли согласно L.Zhivotovsky et al., 2004.
При анализе древней ДНК методом гнездовой ПЦР амплифицировали три перекрывающихся фрагмента ГВа 15986-16149, 16106-16271 и 16209-16401. Продукты амплификации разделяли электрофорезом в 3%-легкоплавкой агарозе, элюировали из геля фрагменты ДНК соответствующей длины с последующей очисткой и переосаждением в этаноле. Полученные фрагменты ДНК секвенировали с обеих цепей с прямыми и обратными праймерами (Yang et al., 1999). Фрагменты ГВСТ мтДНК, результаты секвенирования которых были неоднозначны, клонировали с использованием набора InsT/Aclone PCR Product Cloning Kit, Fermentas. Для каждого фрагмента анализировали несколько бактериальных клонов, содержащих вставку соответствующей длины.
Генотипирование локусов, содержащих Alu-инсерционные полиморфизмы (ACE, APOA1, PV92, TPA25, Ya5NBC27, Ya5NBC102, Ya5NBC148, Ya5NBC182) проводили согласно опубликованным ранее протоколам (Arcot et al. 1996, 1997, 1998; Stoneking et al. 1997; Caroll et al., 2001).
Анализ числа CTG-повторов DMPK гена проводили с применением праймеров, описанных ранее (Brook et al., 1992). Генотипирование Cys282Tyr и His63Asp мутаций гена HFE проводилось по соответствующим протоколам (Beutler et al., 1996).
Статистический анализ базировался на частотах линий и гаплогрупп мтДНК и Y-хромосомы. 95%-ные доверительные интервалы для частот гаплогрупп рассчитывали в программе Sampling, любезно предоставленной V.Macaulay и адаптированной М.Мetspalu (Эстонский Биоцентр). Для оценки генетических взаимоотношений между популяциями использовали метод главных компонент, кластерный анализ и определение вероятности совпадения линий между популяциями. Для проведения факторного анализа с применением метода главных компонент использовали программу POPSTR, любезно предоставленную H.Harpending (Эстонский Биоцентр), и пакет прикладных программ Statistica, version 5.5. (StatSoft Inc., USA). Для проведения кластерного анализа были использованы матрицы генетических расстояний Fst (Reynolds et al., 1983), рассчитанные в пакете программ ARLEQUIN, version 3.01. Филогенетические деревья строили по методу связывания ближайших соседей (neighbor-joining) с использованием программы MEGA, version 2.1. Вероятность совпадения линий (m) между популяциями определяли по формуле: m=∑xiyi, где xi и yi - частоты совпадающей линии i в популяциях X и Y, соответственно.
Расчет показателей генетического разнообразия (h) (Nei, 1987) и анализ молекулярной дисперсии (AMOVA, Excoffier et al., 1992) проводили в пакете программ ARLEQUIN, version 3.01. Значимость Fst оценивали числом пермутаций 10000. Степень генетической дифференциации между изученными популяциями оценивали также с помощью стандартизированной Gst (Gst'), предложенной P.W.Hedrick (Hedrick, 2005), которая позволяет проводить сравнение между локусами с различной степенью генетической вариабельности и скоростью мутирования.
Тестирование соответствия распределения генотипов закону Харди-Вайнберга по Alu-инсерционным локусам, расчет фактической и теоретической гетерозиготности проводили в программе GenePop, version 3.3. Степень генетической дифференциации между популяциями по Alu-инсерционным локусам оценивали с помощью показателя Gst, который рассчитывали по Нею (Nei, 1983). Анализ потока генов проводили в соответствии с моделью, предложенной KHarpending and R.Ward (1982).
Статистический анализ полученных данных по микросателлитным локусам проводили с использованием компьютерных программ Microsat и GenPop (version 3.3). Для оценки генетических расстояний между популяциями использовали меру Sfi (Goldstein et al., 1995). Тестирование на соответствие распределения частот генотипов закону Харди-Вайнберга проводили по модифицированному методу цепи Маркова (Guo and Thompson, 1992).
РЕЗУЛЬТАТЫ И ОБСУЖДЕНИЕ 1. Структура и происхождение митохондриального генофонда популяций Якутии
1.1. Особенности структуры митохондриального генофонда популяций Якутии. Филогенетическая сеть гаплотипов митохондриальной ДНК в популяциях Якутии (n=694) показана на рис.1.

В суммарной выборке выявлено 126 гаплотипов, относящихся к 17 гаплогруппам. Из них 107 линий (91%) относятся к кластерам A, B, C, D, F, G, M7, M13а, Y, Z, характерным для народов Восточной Евразии, 18 гаплотипов (9%) принадлежат к гаплогруппам H, U, HV1, J, T, W, имеющим широкое распространение в Европе и на Ближнем Востоке. Подавляющее большинство линий мтДНК (82%) в популяциях Якутии относится к азиатскому макрокластеру M (С, D, Z, G, M7, M13а), ограниченное число гаплотипов принадлежит к двум другим базовым евразийским макрогаплогруппам N (4%) и R (14%). Характерной особенностью пула митохондриальной ДНК популяций Якутии в сравнении с другими регионами является преобладание двух гаплогрупп - С и D, широко распространенных в Северной Азии.
Частота гаплогруппы С в Якутии достигает максимальных значений в сравнении с другими регионами мира: она составляет 44% митохондриального пула популяций РС(Я). Линии гаплогруппы С в Якутии за одним исключением относятся к двум основным ветвям - C4 и C5. Спектр линий характеризуется высоким разнообразием. Частота второй мажорной гаплогруппы D варьирует от 14% в популяции юкагиров до 33% в популяции центральных якутов (в среднем 28%). Линии гаплогруппы D относятся к двум кластерам - D4 и D5. Кластер D4 представлен в Якутии разнообразным спектром линий - D4a, D4b, D4c, D4h, D4e (D2b), D4i, D4l, D4o2, широко распространенным в более южных популяциях Дальнего Востока и Китая (Kong et al., 2003; Starikovskaya et al., 2005; Yao et al., 2002a, 2002b, 2002c, 2004). Кластер D5, характерный в большей степени для популяций Китая (Kivisild et al., 2002; Yao et al., 2002a, 2002b, 2002c; Nishimaki et al., 1999; Tsai et al., 2001; Qian et al., 2001), представлен в Якутии небольшим количеством линий ветви D5a2 и единственной линией cубкластера D5b1.
Вклад каждой из остальных 15 гаплогрупп в митохондриальный генофонд популяций Якутии составляет, за редким исключением, менее 5%. Присутствие в Якутии гаплогрупп, характерных для более южных регионов, очевидно, связано с древними миграционными процессами в направлении юг-север, имевшими место в прошлом. Гаплогруппа А, широко распространенная в популяциях Восточной Азии, представлена ограниченным количеством линий А4 и А8 в генофонде центральных и вилюйских якутов, эвенков и долган. Гаплогруппа В, характерная для популяций Юго-Восточной Азии, Южной Сибири и Средней Азии, представлена в Якутии единичными линиями субкластеров В4а, B4b1 и B5b2. Гаплогруппа F, также как и В, имеющая южноазиатское происхождение (Ballinger et al., 2002), представлена двумя субкластерами - F1b и F2a. Субкластер F1b наиболее часто встречается в Китае, Южной и Западной Сибири, Средней Азии (Yao et al., 2002a, 2002b, 2002c; Qian et al., 2001; Tsai et al., 2001; Nishimaki et al., 1999; Derenko et al., 2003; Straikovskaya et al., 2005; Comas et al., 1998, 2004; Quintana-Murci et al., 2004; Yao et al., 2004), F2a более характерен для популяций Китая (Yao et al., 2002a, 2002b, 2002c; Qian et al., 2001; Nishimaki et al., 1999). Гаплогруппа М7, характерная для популяций Китая, Кореи, Японии, Южной Сибири представлена в Якутии единичными линиями M7a, M7b, М7с и M7d. Гаплогруппа Y, регионом возникновения которой считается Дальний Восток (Schurr et al., 1999), в Якутии имеет низкую частоту встречаемости (в среднем 1.4%) в популяциях эвенов, центральных и вилюйских якутов. Гаплогруппа G2a, которая распространилась в другие регионы предположительно из Средней Азии (Comas et al., 2004), имеет более высокие частоты в популяциях якутов.
В то же время, результаты анализа линий мтДНК указывают на то, что поток генов в Якутию с восточных регионов Чукотки и Камчатки был ограничен. В популяциях Якутии не выявлены линии субкластера А2, который является преобладающим компонентом митохондриального генофонда чукчей и эскимосов (68% и 77% соответственно) (Starikovskaya et al., 1998). Гаплогруппа G1b, специфичная для популяций побережья Тихого океана - нивхов, ульчей, негидальцев (Starikovskaya et al., 2005), ороков (Бермишева, 2005), коряков, ительменов и чукчей (Starikovskaya et al., 1998; Schurr et al.,1999), представлена в митохондриальном пуле популяций Якутии единичными линиями и имеет максимальную частоту в популяции эвенов (8%). Более высокие частоты в северных популяциях Якутии имеет гаплогруппа Z, но Z1-гаплотипы эвенов и юкагиров Якутии не содержат транзиции 11078, характерной для Z1-линий коряков (Schurr et al., 1999). Этот факт указывает на то, что происхождение Z1 в популяциях Якутии не может быть объяснено проникновением этих линий с территории Камчатки и, скорее всего, связано с более южными регионами.
Распределение частот митохондриальных гаплогрупп в популяциях Якутии в целом имеет сходный характер. Своеобразие митохондриальных портретов проявляется в отношении спектра субгаплогрупп и минорных гаплогрупп (рис.2А).
Частота гаплогруппы С в Якутии достигает максимальных значений в сравнении с другими регионами мира: она составляет 44% митохондриального пула популяций РС(Я). Линии гаплогруппы С в Якутии за одним исключением относятся к двум основным ветвям - C4 и C5. Спектр линий характеризуется высоким разнообразием. Частота второй мажорной гаплогруппы D варьирует от 14% в популяции юкагиров до 33% в популяции центральных якутов (в среднем 28%). Линии гаплогруппы D относятся к двум кластерам - D4 и D5. Кластер D4 представлен в Якутии разнообразным спектром линий - D4a, D4b, D4c, D4h, D4e (D2b), D4i, D4l, D4o2, широко распространенным в более южных популяциях Дальнего Востока и Китая (Kong et al., 2003; Starikovskaya et al., 2005; Yao et al., 2002a, 2002b, 2002c, 2004). Кластер D5, характерный в большей степени для популяций Китая (Kivisild et al., 2002; Yao et al., 2002a, 2002b, 2002c; Nishimaki et al., 1999; Tsai et al., 2001; Qian et al., 2001), представлен в Якутии небольшим количеством линий ветви D5a2 и единственной линией cубкластера D5b1.
Вклад каждой из остальных 15 гаплогрупп в митохондриальный генофонд популяций Якутии составляет, за редким исключением, менее 5%. Присутствие в Якутии гаплогрупп, характерных для более южных регионов, очевидно, связано с древними миграционными процессами в направлении юг-север, имевшими место в прошлом. Гаплогруппа А, широко распространенная в популяциях Восточной Азии, представлена ограниченным количеством линий А4 и А8 в генофонде центральных и вилюйских якутов, эвенков и долган. Гаплогруппа В, характерная для популяций Юго-Восточной Азии, Южной Сибири и Средней Азии, представлена в Якутии единичными линиями субкластеров В4а, B4b1 и B5b2. Гаплогруппа F, также как и В, имеющая южноазиатское происхождение (Ballinger et al., 2002), представлена двумя субкластерами - F1b и F2a. Субкластер F1b наиболее часто встречается в Китае, Южной и Западной Сибири, Средней Азии (Yao et al., 2002a, 2002b, 2002c; Qian et al., 2001; Tsai et al., 2001; Nishimaki et al., 1999; Derenko et al., 2003; Straikovskaya et al., 2005; Comas et al., 1998, 2004; Quintana-Murci et al., 2004; Yao et al., 2004), F2a более характерен для популяций Китая (Yao et al., 2002a, 2002b, 2002c; Qian et al., 2001; Nishimaki et al., 1999). Гаплогруппа М7, характерная для популяций Китая, Кореи, Японии, Южной Сибири представлена в Якутии единичными линиями M7a, M7b, М7с и M7d. Гаплогруппа Y, регионом возникновения которой считается Дальний Восток (Schurr et al., 1999), в Якутии имеет низкую частоту встречаемости (в среднем 1.4%) в популяциях эвенов, центральных и вилюйских якутов. Гаплогруппа G2a, которая распространилась в другие регионы предположительно из Средней Азии (Comas et al., 2004), имеет более высокие частоты в популяциях якутов.
В то же время, результаты анализа линий мтДНК указывают на то, что поток генов в Якутию с восточных регионов Чукотки и Камчатки был ограничен. В популяциях Якутии не выявлены линии субкластера А2, который является преобладающим компонентом митохондриального генофонда чукчей и эскимосов (68% и 77% соответственно) (Starikovskaya et al., 1998). Гаплогруппа G1b, специфичная для популяций побережья Тихого океана - нивхов, ульчей, негидальцев (Starikovskaya et al., 2005), ороков (Бермишева, 2005), коряков, ительменов и чукчей (Starikovskaya et al., 1998; Schurr et al.,1999), представлена в митохондриальном пуле популяций Якутии единичными линиями и имеет максимальную частоту в популяции эвенов (8%). Более высокие частоты в северных популяциях Якутии имеет гаплогруппа Z, но Z1-гаплотипы эвенов и юкагиров Якутии не содержат транзиции 11078, характерной для Z1-линий коряков (Schurr et al., 1999). Этот факт указывает на то, что происхождение Z1 в популяциях Якутии не может быть объяснено проникновением этих линий с территории Камчатки и, скорее всего, связано с более южными регионами.
Распределение частот митохондриальных гаплогрупп в популяциях Якутии в целом имеет сходный характер. Своеобразие митохондриальных портретов проявляется в отношении спектра субгаплогрупп и минорных гаплогрупп (рис.2А).
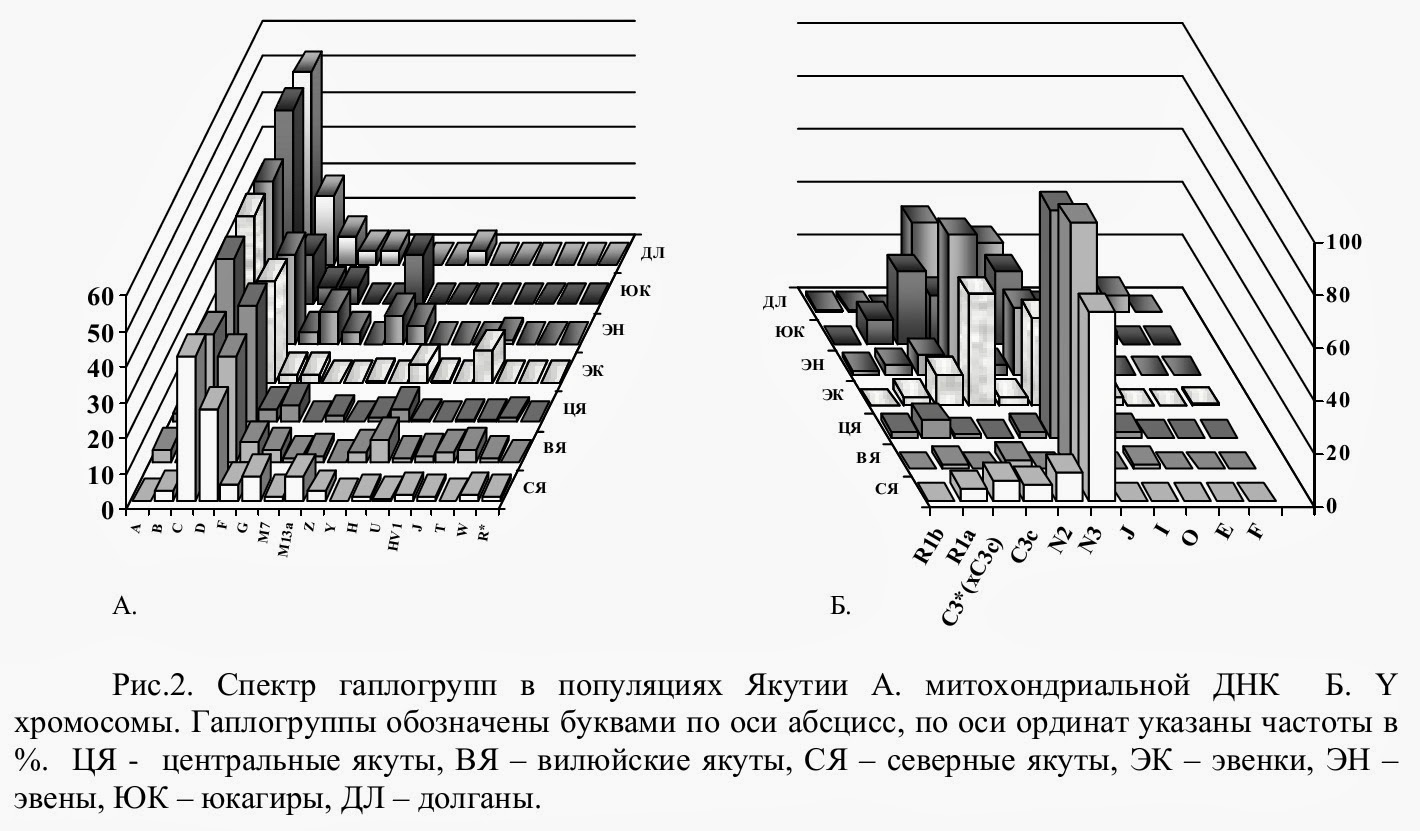
Линии Z1 и G1b больше представлены в популяциях эвенов и юкагиров. Эти гаплогруппы практически отсутствуют в центральной части Якутии - у якутов Лено-Амгинского междуречья и Вилюя. Частота гаплогруппы J у эвенков достоверно выше, чем в других изученных популяциях Якутии за исключением вилюйских якутов. Митохондриальный пул якутов, проживающих в бассейне средней Лены, Амги и Вилюя, отличается достоверно высокой частотой гаплогруппы D5а2. Северные якуты характеризуются более высоким содержанием гаплогрупп B4, Z и М13а.
Уровень генетического разнообразия линий мтДНК в популяциях Якутии ниже, чем в популяциях Средней Азии и Южной Сибири (за исключением сойотов и тофаларов), но выше, чем в популяциях малочисленных этносов Западной Сибири (кроме манси), Дальнего Востока, Чукотки и Камчатки. Из 126 гаплотипов, выявленных в изученной выборке, 63 гаплотипа совпадает между двумя или более
популяциями. Частоты популяционно-специфичных линий варьируют от 8.7% у северных якутов до 22% у эвенков.
1.2. Западноевразийские линии встречаются в генофонде народов Якутии с низкой частотой - их суммарное содержание в общей выборке составляет ~ 9%. Содержание западноевразийских линий минимально в митохондриальном генофонде эвенов (1%) и юкагиров (0%). Вклад линий, распространенных в европейских популяциях, выше в мт-генофонде эвенков (15%) и якутов (в среднем 10%) (рис.3 А). Спектр западноевразийских гаплотипов характеризуется большим разнообразием в популяциях якутов (H, U, HV1, T, W, J). В популяции эвенков выявлено три западноевразийские гаплогруппы - H, U и J.
Чтобы определить происхождение западноевразийских линий, обнаруженных в популяциях Якутии, мы провели их поиск в популяциях соседних регионов (Южная Сибирь и Монголия, Западная Сибирь) в суммарной выборке из 1248 человек, а также в различных популяциях русских (Malyarchuk and Derenko, 2001; Малярчук, 2002; Malyarchuk et al., 2004; Orekhov et al., 1999; Belyaeva et al., 2003) и других европейских этносов - поляков, латвийцев, башкир, татар, чувашей, мордвы, марийцев, удмуртов и коми (Malyarchuk et al., 2002; Pliss et al., 2005; Бермишева, 2002) общей численностью 2657 индивидов. Только 8 из 17 западноевразийских линий популяций Якутии совпадает с линиями русских, 5 - с популяциями Южной Сибири (тувинцами, бурятами, тофаларами), 2 - с популяциями Западной Сибири (манси, кеты, нганасаны).
Анализ совпадающих линий между популяциями показывает, что присутствие европеоидного компонента в генофонде коренного населения РС(Я) может объясняться не только смешением с русскими и другими европейскими этносами. Часть этих линий, очевидно, имеет иное, более древнее происхождение, не связанное с недавней, в масштабах эволюционного времени, миграцией русскоязычного населения в Восточную Сибирь начиная с XVII в. Из исторических данных известно, что численность женщин русской национальности в Якутии в XVII-XVIII в. была крайне низкой (Фишер, 1774). Русское население в Якутии в XVII в. составляли главным образом мужчины, которые женились на местных женщинах. Только с конца XVIII в. началось более массовое переселение в Якутскую губернию семей крестьян-земледельцев и почтовых работников, обслуживающих ямщицкие тракты (Сафронов, 1987). Численность русского населения росла до XX вв. очень медленно: по данным Первой всеобщей переписи российской империи 1897 г. русские Якутской области составляли 30807 человек, или 11.4% от общей численности населения (Романов, 1998).
Уровень генетического разнообразия линий мтДНК в популяциях Якутии ниже, чем в популяциях Средней Азии и Южной Сибири (за исключением сойотов и тофаларов), но выше, чем в популяциях малочисленных этносов Западной Сибири (кроме манси), Дальнего Востока, Чукотки и Камчатки. Из 126 гаплотипов, выявленных в изученной выборке, 63 гаплотипа совпадает между двумя или более
популяциями. Частоты популяционно-специфичных линий варьируют от 8.7% у северных якутов до 22% у эвенков.
1.2. Западноевразийские линии встречаются в генофонде народов Якутии с низкой частотой - их суммарное содержание в общей выборке составляет ~ 9%. Содержание западноевразийских линий минимально в митохондриальном генофонде эвенов (1%) и юкагиров (0%). Вклад линий, распространенных в европейских популяциях, выше в мт-генофонде эвенков (15%) и якутов (в среднем 10%) (рис.3 А). Спектр западноевразийских гаплотипов характеризуется большим разнообразием в популяциях якутов (H, U, HV1, T, W, J). В популяции эвенков выявлено три западноевразийские гаплогруппы - H, U и J.
Чтобы определить происхождение западноевразийских линий, обнаруженных в популяциях Якутии, мы провели их поиск в популяциях соседних регионов (Южная Сибирь и Монголия, Западная Сибирь) в суммарной выборке из 1248 человек, а также в различных популяциях русских (Malyarchuk and Derenko, 2001; Малярчук, 2002; Malyarchuk et al., 2004; Orekhov et al., 1999; Belyaeva et al., 2003) и других европейских этносов - поляков, латвийцев, башкир, татар, чувашей, мордвы, марийцев, удмуртов и коми (Malyarchuk et al., 2002; Pliss et al., 2005; Бермишева, 2002) общей численностью 2657 индивидов. Только 8 из 17 западноевразийских линий популяций Якутии совпадает с линиями русских, 5 - с популяциями Южной Сибири (тувинцами, бурятами, тофаларами), 2 - с популяциями Западной Сибири (манси, кеты, нганасаны).
Анализ совпадающих линий между популяциями показывает, что присутствие европеоидного компонента в генофонде коренного населения РС(Я) может объясняться не только смешением с русскими и другими европейскими этносами. Часть этих линий, очевидно, имеет иное, более древнее происхождение, не связанное с недавней, в масштабах эволюционного времени, миграцией русскоязычного населения в Восточную Сибирь начиная с XVII в. Из исторических данных известно, что численность женщин русской национальности в Якутии в XVII-XVIII в. была крайне низкой (Фишер, 1774). Русское население в Якутии в XVII в. составляли главным образом мужчины, которые женились на местных женщинах. Только с конца XVIII в. началось более массовое переселение в Якутскую губернию семей крестьян-земледельцев и почтовых работников, обслуживающих ямщицкие тракты (Сафронов, 1987). Численность русского населения росла до XX вв. очень медленно: по данным Первой всеобщей переписи российской империи 1897 г. русские Якутской области составляли 30807 человек, или 11.4% от общей численности населения (Романов, 1998).

Вероятным источником некоторых западноевразийских линий в генофонде популяций Якутии может быть древнее население Южной Сибири. Гаплогруппы H, J и U представляют собой основной европеоидный компонент генофондов народов Южной Сибири и Средней Азии (Derenko et al., 2003; Starikovskaya et al., 2005; Comas et al.,1998, 2004; Quintana-Murci, 2004). Гаплогруппы Н и U4 также характерны для популяций Западной Сибири - кетов, манси, нганасанов (Дербенева, 2002; Derbeneva et al., 2002; Малярчук, 2004). Не исключено, что происхождение некоторых из Н и U4 линий в Якутии может быть связано с древним палеоевропеоидным уралоязычным населением Западной Сибири.
1.3. Филогеография мажорных гаплогрупп С и Б5а2.
Гаплогруппа С. Гаплогруппа С типична для популяций Сибири, поэтому наиболее вероятным представляется ее возникновение в одном из регионов Северо-Востока Евразии. Для гаплогруппы С были оценены частоты, время коалесценции и число линий в различных регионах (Чукотка и Камчатка, Дальний Восток, Китай, Южная Сибирь и Монголия, Западная Сибирь и Средняя Азия) (табл.2).

Общее время коалесценции линий находится в интервале 39500±5300 лет. Высокая частота и разнообразие линий гаплогруппы С, присутствие линий всех основных ветвей (С1, С4а1, С4а2, С4Ь1а и С5), наибольшее количество специфичных линий характерны для популяций Южной Сибири и Монголии, что позволяет предположить, что именно этот регион является наиболее вероятным местом возникновения HgC, откуда последовала экспансия ее носителей по всей территории северо-востока Евразии и дальнейшая дивергенция уже в условиях региональной изоляции.
Филогенетическое дерево гаплогруппы С показано на рис.4.
По-видимому, Якутия была главным транзитным регионом при миграции носителей гаплогруппы С на Чукотку и Камчатку. Популяции Чукотки и Камчатки характеризуются исключительно высокой частотой специфичных С-линий: три четверти гаплотипов образуют два регионально-специфических кластера, один из которых (С4b2) определяется транзицией в позиции 16124, другой, относящийся к С5а, сочетанием мутаций 16093-16189-16223-16261-16288-16298 (рис.4). Наличие двух специфических субкластеров, имеющих высокую степень дивергенции, указывает на длительную эволюцию популяций данного региона в изолированных условиях. Время коалесценции линий первого субкластера - 5000±2500 и 7900±2600 - для второго. Если принять во внимание минимальное значение эволюционного времени, необходимого для возникновения рассматриваемых кластеров, то можно сделать заключение, что в течение по крайней мере последних 2500-5300 лет поток генов между популяциями Чукотки и Камчатки и популяциями соседних регионов, в том числе Якутии, был крайне ограничен.
В филогенетической сети гаплогруппы C не выявлено кластеров с высоким уровнем дивергенции, которые были бы характерны только для Якутии (в отличие от регионов Чукотки, Южной Сибири и Китая), хотя более трети С-линий популяций РС(Я) являются специфичными. Этот факт указывает на постоянные связи населения Якутии с соседними регионами и соответствует мнению историков и археологов, что вся этническая история Якутии насыщена многочисленными свидетельствами культурных связей с другими народами Сибири, начиная с палеолита и до наших дней (Мочанов, 1993; Алексеев, 1996a).
Гаплогруппа D5a2. В Якутии широко представлен субкластер D5a2, время коалесценции которого оценено в 55700±15500 лет. Субкластер D5а2, по всей видимости, возник в Китае: высокая частота и разнообразие D5a2-линий наблюдаются в популяциях Китая (табл.2).С территории Китая произошло распространение D5a2-носителей в более северные регионы Южной Сибири, Монголии и Дальнего Востока. На рис.5 показана филогенетическая сеть линий кластера D5a2.

1.4. Генетические взаимоотношения между популяциями Якутии. Для определения генетических взаимоотношений между популяциями были использованы три подхода: метод главных компонент, базирующийся на частотах гаплогрупп в популяциях, оценка количества совпадающих линий и филогенетический анализ, базирующийся на Fst-расстояниях между популяциями (Reynolds et al., 1983).
В пространстве двух главных компонент, описывающих 61% вариабельности гаплогрупп мтДНК, популяции Якутии образуют три кластера: первый включает популяции юкагиров и эвенов, второй - популяции якутов, третий кластер образован эвенками и долганами (рис.6А).

По первой компоненте, описывающей 41.7% наблюдаемой вариабельности мт-гаплогрупп, юкагиры и эвены отделены от других популяций вследствие преобладания гаплогрупп Z, G1, B и отсутствия западноевразийских гаплогрупп. Отличия популяций якутов от эвенков и долган по второй компоненте определяются главным образом различным соотношением западноевразийских гаплогрупп (W, HV1, T, U против J, H) и восточноевразийских кластеров (G2, D5 против A) в их митохондриальном генофонде.
Максимальные вероятности совпадения гаплотипов мтДНК выявлены между популяциями центральных и вилюйских якутов, и между эвенами и юкагирами. Высокая степень совпадения гаплотипов наблюдается между всеми популяциями якутов и эвенками, также между эвенками, эвенами и долганами: в большинстве случаев количество совпадающих линий между этими популяциями варьирует от 50% до 66%. Наиболее низкая вероятность совпадения линий мтДНК обнаружена между вилюйскими якутами и юкагирами.
Дендрограмма генетических взимоотношений между популяциями Якутии, построенная по методу связывания ближайших соседей на основе матрицы Fst расстояний демонстрирует отдаленность юкагиров и эвенов от остальных популяций Якутии и близкое генетическое родство центральных и вилюйских якутов (рис.бБ).
В целом, результаты оценки генетических взаимоотношений между популяциями с использованием всех трех подходов свидетельствуют о тесном генетическом родстве между центральными и вилюйскими якутами, а также между юкагирами и эвенами. Максимальные различия выявляются между митохондриальными генофондами вилюйских якутов и юкагиров.
1.5. Результаты AMOVA. Когда все семь популяций рассматриваются как одна группа, 98.2% генетической вариабельности линий мтДНК определяются внутри-популяционными различиями и 1.8% межпопуляционными. По частотам мт-гаплогрупп степень генетических различий между популяциями имеет еще более низкое значение - 0.9%.
Следует отметить, что степень генетической подразделенности популяций Якутии ниже, чем во всех остальных регионах Сибири (Чукотка и Камчатка - 0.333, Дальний Восток и Северный Китай - 0.182, Южная Сибирь - 0.061, Западная Сибирь - 0.046) (табл.3) и в Средней Азии (2.34%) (Comas et al., 2004), несмотря на значительные географические расстояния между популяциями Якутии.
Низкая степень генетических различий между популяциями Якутии указывает на их общее происхождение и/ или достаточно интенсивный поток генов между ними.

1.6. Генетические взаимоотношения между популяциями Якутии и популяциями соседних регионов. Для оценки генетических взаимоотношений между популяциями были использованы три подхода: факторный анализ, определение вероятности совпадения линий между популяциями (m) и кластерный анализ. В пространстве двух главных компонент популяции делятся на три кластера (рис.7).
.jpg)
Первый, достаточно компактный кластер образован популяциями Южной и Западной Сибири, в него также входят эвенки Северного Китая и орочоны, во второй вошли популяции Дальнего Востока, коряки и ительмены, третий, достаточно удаленный от первых двух кластер включает чукчей и эскимосов. Популяции Якутии входят в первый кластер, образованный в основном популяциями Южной и Западной Сибири.
Наибольшая вероятность совпадения линий обнаружена для всех популяций РС(Я) с популяциями Южной Сибири, затем, по мере уменьшения, с популяциями Западной Сибири, Северного Китая и Дальнего Востока и, наконец, Средней Азии, Чукотки и Камчатки.
Кластерный анализ проводили на основе матрицы Fst дистанций (Reynolds et al., 1983). Изученные популяции Якутии входят в один кластер с популяциями Южной Сибири (тувинцами, тофаларами) и западными эвенками, удалены от популяций Дальнего Востока и Северного Китая, Чукотки и Камчатки.
В целом, согласно полученным результатам, по составу линий митохондриальной ДНК популяции Якутии наиболее близки к популяциям Южной Сибири. Из соседних регионов наибольшие различия по мт-линиям у популяций Якутии выявлены с популяциями Чукотки и Камчатки.
2.Анализ линий древней митохондриальной ДНК в Якутии
Исследована митохондриальная ДНК из костных останков могильников Якутии с целью идентификации, типирования и проведения сравнительного анализа с линиями современных популяций. Выявлено 5 различных гаплотипов мтДНК, характерных для популяций Азии (табл.4).
Наибольшая вероятность совпадения линий обнаружена для всех популяций РС(Я) с популяциями Южной Сибири, затем, по мере уменьшения, с популяциями Западной Сибири, Северного Китая и Дальнего Востока и, наконец, Средней Азии, Чукотки и Камчатки.
Кластерный анализ проводили на основе матрицы Fst дистанций (Reynolds et al., 1983). Изученные популяции Якутии входят в один кластер с популяциями Южной Сибири (тувинцами, тофаларами) и западными эвенками, удалены от популяций Дальнего Востока и Северного Китая, Чукотки и Камчатки.
В целом, согласно полученным результатам, по составу линий митохондриальной ДНК популяции Якутии наиболее близки к популяциям Южной Сибири. Из соседних регионов наибольшие различия по мт-линиям у популяций Якутии выявлены с популяциями Чукотки и Камчатки.
2.Анализ линий древней митохондриальной ДНК в Якутии
Исследована митохондриальная ДНК из костных останков могильников Якутии с целью идентификации, типирования и проведения сравнительного анализа с линиями современных популяций. Выявлено 5 различных гаплотипов мтДНК, характерных для популяций Азии (табл.4).

Проведен поиск этих типов мтДНК в базе данных с линиями более 40000 человек из современных популяций мира, в том числе 900 из Якутии (наши данные; Пузырев, 2003; Pakendorf et al., 2003).
2.1.Гаплотипы мтДНК якутов XVIII в. Линии субкластера С4а, к которому относится гаплотип образца №1 из Ат-Дабаанского погребения, типичны для популяций Сибири (Derenko et al., 2003; Starikovskaya et al., 2005; Derbeneva et al., 2002; Дербенева, 2002), но редко встречаются в Средней Азии и Китае (Tsai et al., 2001; Kivisild et al., 2002; Yao et al., 2002a, 2002b, 2002c, 2004; Nishimaki et al., 1999; Comas et al., 1998). Присутствие линии индивида из Ат-Дабаанского погребения 16223-16298-16327-16344-16357 в современных популяциях якутов, тувинцев, тофаларов, алтайцев указывает на древние генетические связи между регионами Южной Сибири и Якутии и соответствует традиционным взглядам о прибайкальском происхождении южных предков якутов.
Линия образца №2 из Одьулуунского относится к субкластеру D5a2, наиболее характерному для митохондриального генофонда якутов. В популяциях центральных и вилюйских якутов частота D5a2 достигает 18%. Следует отметить, что линии D5a2 обнаружены также в 5 образцах древней ДНК из якутских погребений в местности Ат-Дабаан (Ricaut et al., 2004; 2006) центральной Якутии.
Полученные результаты указывают на преемственность гаплотипов образцов №1 и №2 в генофонде якутов за последние 300 лет.
Гаплотип третьего индивида из Дьарааминского погребения характеризуется полиморфизмами 16140-16189-16243, типичными для ветви В5b гаплогруппы В. В изученной нами выборке субкластер B5b представлен единственной линией 1611116140-16189-16234-16243, найденной у одного якута Хангаласского улуса, отличающейся на две мутации от линии образца №3 древней ДНК. Возможно, что низкая частота B5b-линий в предковой популяции якутов способствовали исчезновению гаплотипа 16111-16140-16189-16223-16243 вследствие эффекта генетического дрейфа, оказывающего сильное влияние на популяции с малой эффективной численностью. Тем не менее, присутствие линий B5b в современной и древней популяциях якутов указывает на их характерность для митохондриального генофонда якутов в течение последних столетий.
2.2. Гаплотипы мтДНК двух человек из Кёрдюгенского погребения эпохи позднего неолита Якутии В Кёрдюгенском погребении эпохи позднего неолита Якутии найдена двойная могила. Линия образца №4 относится к гаплогруппе А4. Любопытно, что в базе данных эта линия обнаружена только в популяции казахов (Comas et al., 1998). Филогенетически близкий гаплотип, отличающийся на 1 мутацию 16039-16189-16223-16290-16319-16356-16362, выявлен в популяциях центральных и вилюйских якутов, у эвенков РС(Я), Средней Сибири и побережья Охотского моря (Torroni et al., 1993; Starikovskaya et al., 2005), манси Западной Сибири (Derbeneva et al., 2002), орочонов Дальнего Востока (Kong et al., 2003).
Гаплотип 16189-16223-16278-16362 образца № 5 Кёрдюгенского погребения не может быть типирован однозначно по полиморфизмам в ГВС1 Позиции 16189, 16278, 16362 являются быстро мутирующими сайтами ГВС1 и встречаются в различных ветвях филогенетического дерева мтДНК. Гаплотип №5 может быть отнесен к гаплогруппам D, G2а, E, которые характеризуются сочетанием полиморфизмов 16223 и 16362. Такой же гаплотип обнаружен у двух монголов (Kong et al., 2003), одного корейца (Kong et al., 2003) и одного бай (Yao et al., 2002a) и классифицирован как относящийся к субкластеру G2(а). Более вероятным представляется принадлежность гаплотипа №5 к кластерам G2а или D, чем к достаточно редкой гаплогруппе Е. Анализ мтДНК позволяет заключить, что два человека, захороненных в одной могиле, не были родственниками по материнской линии.
Мнения исследователей относительно этнической принадлежности поздненеолитических племен ымыяхтахцев неоднозначны: некоторые связывают их с предками юкагиров (Левин, 1958; Симченко, 1976), другие считают их прародителями чукчей, эскимосов и коряков (Константинов, 1978; Диков, 1979). По мнению признанного исследователя ымыяхтахской культуры С. А. Федосеевой, ымыяхтахцы могли участвовать в формировании нескольких современных этносов -юкагиров и северо-восточных палеоазиатов (Федосеева, 1980). Это мнение поддерживается А.Н.Алексеевым (Алексеев, 1996a) исходя из того, что в ымыяхтахской культуре выделяются несколько локальных вариантов, за которыми могут стоять различные этнические образования, а также на основе краниологического анализа поздненеолитических останков человека.
В Якутии известно несколько погребений позднего неолита: это могильники Чочур-Муран и Диринг-Юрях на Лене, погребения Родинка, Помазкино и Каменка (на Колыме), Куллаты, Иччилях, Вилюйское шоссе (на Лене), Кёрдюген (Алексеев, 2006; Окладников, 1955a; Федосеева, 1980; Федосеева, 1992; Гохман и Томтосова, 1992; Кистенев, 1992; Кашин, 2001; Шпакова, 2001; Дьяконов, 2003). Серия черепов шести человек погребения Диринг-Юрях имела своеобразные морфологические особенности, не характерные для современных монголоидов. Такие особенности были найдены в отдельных группах арктической и центральноазиатской рас. При сравнении диринг-юряхских черепов с краниологическими сериями современных народов Восточной Сибири, наибольшее сходство было обнаружено с якутами (Гохман и Томтосова, 1992).Отсутствие достаточного количества изученных погребений и антропологических данных эпохи позднего неолита не позволяет однозначно определить генетические связи ымыяхтахского населения Якутии с современными этносами. Из молекулярно-генетических работ до сих пор известна лишь одна публикация F.-X. Ricaut и соавт. (2005), в которой анализировали мтДНК неолитической женщины из Родинкского погребения. Установленный гаплотип 16223-16298-16327 относится к гаплогруппе С. Это женское погребение эпохи позднего неолита обнаружено в Северо-Восточной Якутии на р.Пантелеихе - правом притоке нижней Колымы - в 1980 г. (Кистенев, 1992) и датировано радиоуглеродным методом 3600±60 лет (Ricaut et al., 2005). Гаплотип ГВО 1622316298-16327 - наиболее древний для гаплогруппы С, широко распространен в популяциях Якутии, Южной и Западной Сибири, Чукотки и Камчатки, Дальнего Востока, Китая, Средней Азии и Америки. Отсутствие специфичности этого гаплотипа не позволяет связать его с каким-либо конкретным современным этносом.
Гаплотипы Кёрдюгенского погребения, сравнительно-типологическим методом датированного временем позднего неолита Якутии, не найдены в современных популяциях Чукотки и Камчатки у чукчей, эскимосов, коряков и ительменов (Starikovskaya et al., 1998; Schurr et al., 1999). Полученные результаты не соответствуют выдвинутым ранее гипотезам о генетической связи ымыяхтахцев с современными палеоазиатскими этносами. Результаты нашей работы больше соответствуют антропологическим исследованиям диринг-юряхской серии черепов позднего неолита (Гохман и Томтосова, 1992) и указывают на то, что происхождение ымыяхтахской культуры, возможно, связано с центральноазиатским регионом. Современные популяции Якутии генетически наиболее близки к популяциям Южной Сибири, и отдалены от популяций Чукотки и Камчатки. Дистанцированность населения Якутии от предков современных палеоазиатов была, по-видимому, характерна и для эпохи позднего неолита.
3. Происхождение и особенности спектра линий Y хромосомы в популяциях Якутии
3.1. Анализ линий Y хромосомы в популяциях Якутии. Филогенетическое дерево линий Y хромосомы, построенное по 26 диагностическим маркерам, представлено на рис.8.
.jpg)
Из четырех гаплогрупп, широко распространенных в Сибири (N,C,R,Q) (Karafet et al., 2002), львиную долю пула Y хромосом в популяциях Якутии составляют две гаплогруппы - N (57.5%) и C (30.5%). Гораздо более низкое содержание линий наблюдается в отношении гаплогруппы R (7%). Линии гаплогрупп J, I, F, E3b, О, Q, G выявлены в популяциях Якутии с низкими частотами: их суммарное содержание в пуле Y хромосом составляет всего 5%.
В популяциях якутов выявлено крайне низкое разнообразие линий Y хромосомы, что соответствует опубликованным ранее данным (Zerjal et al., 1997; Pakendorf et al., 2002; Степанов, 2002). Частота мажорной гаплогруппы N3 (N1C) составляет от 71% в популяции северных якутов до 93% у вилюйских якутов. Преобладание ТАТ-С хромосом является характерной чертой для мужского генофонда якутов в целом. Эта особенность резко отличает якутов от других ранее изученных этносов. В популяциях эвенков, эвенов, юкагиров и долган содержание гаплогруппы N3 (N1C) намного ниже. В этих популяциях по частоте доминирует кластер С3 (от 34% у долган до 60% у эвенов) (рис.2Б). Преобладающая часть линий гаплогруппы С3 в популяциях эвенков, эвенов и долган относится к ветви С3с, встречающейся с высокими частотами у народов тунгусо-маньчжурской (эвенки, орочоны, удегейцы, ульчи, негидальцы) (Lell et al., 2002; Karafet et al., 2002) и монгольской (монголы, урянхаи, захчины) (Wells et al., 2001; Katoh et al., 2004) групп алтайской языковой семьи, в популяциях казахов (Karafet et al., 2002; Wells et al., 2001), коряков и ительменов (Lell et al., 2002). С3* линии имеют большую частоту в популяции юкагиров.
Гаплогруппа R представлена в Якутии двумя ветвями R1a и R1b10 (R1b1b2). Линии R1a, встречающиеся с максимальными частотами в популяциях Восточной Европы и Средней Азии (Semino et al., 2000a, Rosser et al., 2000) присутствуют в генофонде всех изученных популяций Якутии. Линии R1b10 (R1b1b2), более характерные для Западной Европы и Ближнего Востока (Semino et al., 2000a; Al-Zahery et al., 2003), обнаружены в популяциях центральных якутов, эвенов и долган.
Гаплогруппа N2 (N1B), частота которой достигает максимума в популяциях некоторых уралоязычных этносов Западной Сибири (Karafet et al., 2002), более представлена в северных широтах Якутии - в популяциях северных якутов, эвенков, эвенов и долган.
Кластер Q, имеющий наибольшие частоты в Западной Сибири и на Чукотке, обнаружен в популяциях эвенов и юкагиров (Karafet et al., 2002; Pakendorf et al., 2006). Другая редкая для Якутии гаплогруппа О, характерная для популяций Юго-Восточной Азии, выявлена в пуле Y-хромосом юкагиров и долган.
3.2. Западноевразийские линии Y хромосомы в популяциях РС(Я). На рис.3Б показано процентное соотношение западно- (G, E, I, J, R1) и восточноевразийских (C, Q, О, N) линий Y хромосомы в изученных популяциях. Гаплогруппа R1 рассматривается нами как западноевразийская условно, принимая во внимание то, что максимальные частоты R1 наблюдаются в восточнославянских популяциях Европы (Semino et al., 2000a, Rosser et al., 2000). Наибольшее содержание западноевразийских линий Y хромосомы обнаружено в популяции долган (26%), наименьшее - у северных и вилюйских якутов (менее 5%). Гаплогруппы R1a и I встречаются в генофонде всех коренных этносов Якутии, тогда как гаплогруппы G, R1b, J, E3b выявлены в отдельных популяциях. Происхождение линий гаплогрупп I, G, E, типичных для Европы и Ближнего Востока, в генофонде коренных народов Якутии, скорее всего, связано с процессами метисации с русскими и, возможно, другими европейскими этносами, заселившими территорию Якутии начиная с XVII в. Линии гаплогруппы I, присутствующие во всех популяциях Якутии, с высокими частотами обнаружены практически во всех восточноевропейских популяциях, не обнаружены в Западной Сибири (за исключением популяций западных эвенков и ненцев) (Karafet et al., 2002; Wells et al., 2001), с низкой частотой встречаются в некоторых популяциях Южной Сибири (Derenko et al., 2006). Также очевидно недавнее проникновение линий гаплогрупп E3b и G в генофонд долган, самого молодого этноса Сибири, сложившегося в период летописной истории с XVII по XIX вв. Согласно данным переписей и исторических документов в состав долган вошли линии русских старожилов Сибири (Долгих, 1963). Гаплогруппа E3b не обнаружена в других популяциях Якутии, Южной и Западной Сибири (за исключением тувинцев), но присутствует в небольших количествах во всех восточноевропейских популяциях. Повышенная частота гаплогруппы G характерна для целого ряда популяций Кавказа (Nasidze et al., 2004) и Ближнего Востока (Semino et al., 2000; Cinnioglu et al., 2004), из восточноевропейских популяций встречается с небольшими частотами у русских и украинцев.
Что касается гаплогрупп R1 а и J, выявленных в изученных популяциях, то не исключается более древнее происхождение части этих линий из южных регионов Алтае-Саянского нагорья: эти гаплогруппы широко распространены в популяциях Южной Сибири и Монголии (Karafet et al., 2002; Katoh et al., 2004; Derenko et al., 2006).
3.3. Разнообразие микросателлитных гаплотипов Y хромосомы в популяциях Якутии. Компаундные гаплотипы Y хромосомы были определены по шести микросателлитным STR-маркерам (DYS19, DYS388, DYS390, DYS391, DYS392, DYS393) у 317 индивидов. Медианные сети N3 (N1C)- и С3-гаплотипов изученных популяций представлены на рис.9.
Линия образца №2 из Одьулуунского относится к субкластеру D5a2, наиболее характерному для митохондриального генофонда якутов. В популяциях центральных и вилюйских якутов частота D5a2 достигает 18%. Следует отметить, что линии D5a2 обнаружены также в 5 образцах древней ДНК из якутских погребений в местности Ат-Дабаан (Ricaut et al., 2004; 2006) центральной Якутии.
Полученные результаты указывают на преемственность гаплотипов образцов №1 и №2 в генофонде якутов за последние 300 лет.
Гаплотип третьего индивида из Дьарааминского погребения характеризуется полиморфизмами 16140-16189-16243, типичными для ветви В5b гаплогруппы В. В изученной нами выборке субкластер B5b представлен единственной линией 1611116140-16189-16234-16243, найденной у одного якута Хангаласского улуса, отличающейся на две мутации от линии образца №3 древней ДНК. Возможно, что низкая частота B5b-линий в предковой популяции якутов способствовали исчезновению гаплотипа 16111-16140-16189-16223-16243 вследствие эффекта генетического дрейфа, оказывающего сильное влияние на популяции с малой эффективной численностью. Тем не менее, присутствие линий B5b в современной и древней популяциях якутов указывает на их характерность для митохондриального генофонда якутов в течение последних столетий.
2.2. Гаплотипы мтДНК двух человек из Кёрдюгенского погребения эпохи позднего неолита Якутии В Кёрдюгенском погребении эпохи позднего неолита Якутии найдена двойная могила. Линия образца №4 относится к гаплогруппе А4. Любопытно, что в базе данных эта линия обнаружена только в популяции казахов (Comas et al., 1998). Филогенетически близкий гаплотип, отличающийся на 1 мутацию 16039-16189-16223-16290-16319-16356-16362, выявлен в популяциях центральных и вилюйских якутов, у эвенков РС(Я), Средней Сибири и побережья Охотского моря (Torroni et al., 1993; Starikovskaya et al., 2005), манси Западной Сибири (Derbeneva et al., 2002), орочонов Дальнего Востока (Kong et al., 2003).
Гаплотип 16189-16223-16278-16362 образца № 5 Кёрдюгенского погребения не может быть типирован однозначно по полиморфизмам в ГВС1 Позиции 16189, 16278, 16362 являются быстро мутирующими сайтами ГВС1 и встречаются в различных ветвях филогенетического дерева мтДНК. Гаплотип №5 может быть отнесен к гаплогруппам D, G2а, E, которые характеризуются сочетанием полиморфизмов 16223 и 16362. Такой же гаплотип обнаружен у двух монголов (Kong et al., 2003), одного корейца (Kong et al., 2003) и одного бай (Yao et al., 2002a) и классифицирован как относящийся к субкластеру G2(а). Более вероятным представляется принадлежность гаплотипа №5 к кластерам G2а или D, чем к достаточно редкой гаплогруппе Е. Анализ мтДНК позволяет заключить, что два человека, захороненных в одной могиле, не были родственниками по материнской линии.
Мнения исследователей относительно этнической принадлежности поздненеолитических племен ымыяхтахцев неоднозначны: некоторые связывают их с предками юкагиров (Левин, 1958; Симченко, 1976), другие считают их прародителями чукчей, эскимосов и коряков (Константинов, 1978; Диков, 1979). По мнению признанного исследователя ымыяхтахской культуры С. А. Федосеевой, ымыяхтахцы могли участвовать в формировании нескольких современных этносов -юкагиров и северо-восточных палеоазиатов (Федосеева, 1980). Это мнение поддерживается А.Н.Алексеевым (Алексеев, 1996a) исходя из того, что в ымыяхтахской культуре выделяются несколько локальных вариантов, за которыми могут стоять различные этнические образования, а также на основе краниологического анализа поздненеолитических останков человека.
В Якутии известно несколько погребений позднего неолита: это могильники Чочур-Муран и Диринг-Юрях на Лене, погребения Родинка, Помазкино и Каменка (на Колыме), Куллаты, Иччилях, Вилюйское шоссе (на Лене), Кёрдюген (Алексеев, 2006; Окладников, 1955a; Федосеева, 1980; Федосеева, 1992; Гохман и Томтосова, 1992; Кистенев, 1992; Кашин, 2001; Шпакова, 2001; Дьяконов, 2003). Серия черепов шести человек погребения Диринг-Юрях имела своеобразные морфологические особенности, не характерные для современных монголоидов. Такие особенности были найдены в отдельных группах арктической и центральноазиатской рас. При сравнении диринг-юряхских черепов с краниологическими сериями современных народов Восточной Сибири, наибольшее сходство было обнаружено с якутами (Гохман и Томтосова, 1992).Отсутствие достаточного количества изученных погребений и антропологических данных эпохи позднего неолита не позволяет однозначно определить генетические связи ымыяхтахского населения Якутии с современными этносами. Из молекулярно-генетических работ до сих пор известна лишь одна публикация F.-X. Ricaut и соавт. (2005), в которой анализировали мтДНК неолитической женщины из Родинкского погребения. Установленный гаплотип 16223-16298-16327 относится к гаплогруппе С. Это женское погребение эпохи позднего неолита обнаружено в Северо-Восточной Якутии на р.Пантелеихе - правом притоке нижней Колымы - в 1980 г. (Кистенев, 1992) и датировано радиоуглеродным методом 3600±60 лет (Ricaut et al., 2005). Гаплотип ГВО 1622316298-16327 - наиболее древний для гаплогруппы С, широко распространен в популяциях Якутии, Южной и Западной Сибири, Чукотки и Камчатки, Дальнего Востока, Китая, Средней Азии и Америки. Отсутствие специфичности этого гаплотипа не позволяет связать его с каким-либо конкретным современным этносом.
Гаплотипы Кёрдюгенского погребения, сравнительно-типологическим методом датированного временем позднего неолита Якутии, не найдены в современных популяциях Чукотки и Камчатки у чукчей, эскимосов, коряков и ительменов (Starikovskaya et al., 1998; Schurr et al., 1999). Полученные результаты не соответствуют выдвинутым ранее гипотезам о генетической связи ымыяхтахцев с современными палеоазиатскими этносами. Результаты нашей работы больше соответствуют антропологическим исследованиям диринг-юряхской серии черепов позднего неолита (Гохман и Томтосова, 1992) и указывают на то, что происхождение ымыяхтахской культуры, возможно, связано с центральноазиатским регионом. Современные популяции Якутии генетически наиболее близки к популяциям Южной Сибири, и отдалены от популяций Чукотки и Камчатки. Дистанцированность населения Якутии от предков современных палеоазиатов была, по-видимому, характерна и для эпохи позднего неолита.
3. Происхождение и особенности спектра линий Y хромосомы в популяциях Якутии
3.1. Анализ линий Y хромосомы в популяциях Якутии. Филогенетическое дерево линий Y хромосомы, построенное по 26 диагностическим маркерам, представлено на рис.8.
В популяциях якутов выявлено крайне низкое разнообразие линий Y хромосомы, что соответствует опубликованным ранее данным (Zerjal et al., 1997; Pakendorf et al., 2002; Степанов, 2002). Частота мажорной гаплогруппы N3 (N1C) составляет от 71% в популяции северных якутов до 93% у вилюйских якутов. Преобладание ТАТ-С хромосом является характерной чертой для мужского генофонда якутов в целом. Эта особенность резко отличает якутов от других ранее изученных этносов. В популяциях эвенков, эвенов, юкагиров и долган содержание гаплогруппы N3 (N1C) намного ниже. В этих популяциях по частоте доминирует кластер С3 (от 34% у долган до 60% у эвенов) (рис.2Б). Преобладающая часть линий гаплогруппы С3 в популяциях эвенков, эвенов и долган относится к ветви С3с, встречающейся с высокими частотами у народов тунгусо-маньчжурской (эвенки, орочоны, удегейцы, ульчи, негидальцы) (Lell et al., 2002; Karafet et al., 2002) и монгольской (монголы, урянхаи, захчины) (Wells et al., 2001; Katoh et al., 2004) групп алтайской языковой семьи, в популяциях казахов (Karafet et al., 2002; Wells et al., 2001), коряков и ительменов (Lell et al., 2002). С3* линии имеют большую частоту в популяции юкагиров.
Гаплогруппа R представлена в Якутии двумя ветвями R1a и R1b10 (R1b1b2). Линии R1a, встречающиеся с максимальными частотами в популяциях Восточной Европы и Средней Азии (Semino et al., 2000a, Rosser et al., 2000) присутствуют в генофонде всех изученных популяций Якутии. Линии R1b10 (R1b1b2), более характерные для Западной Европы и Ближнего Востока (Semino et al., 2000a; Al-Zahery et al., 2003), обнаружены в популяциях центральных якутов, эвенов и долган.
Гаплогруппа N2 (N1B), частота которой достигает максимума в популяциях некоторых уралоязычных этносов Западной Сибири (Karafet et al., 2002), более представлена в северных широтах Якутии - в популяциях северных якутов, эвенков, эвенов и долган.
Кластер Q, имеющий наибольшие частоты в Западной Сибири и на Чукотке, обнаружен в популяциях эвенов и юкагиров (Karafet et al., 2002; Pakendorf et al., 2006). Другая редкая для Якутии гаплогруппа О, характерная для популяций Юго-Восточной Азии, выявлена в пуле Y-хромосом юкагиров и долган.
3.2. Западноевразийские линии Y хромосомы в популяциях РС(Я). На рис.3Б показано процентное соотношение западно- (G, E, I, J, R1) и восточноевразийских (C, Q, О, N) линий Y хромосомы в изученных популяциях. Гаплогруппа R1 рассматривается нами как западноевразийская условно, принимая во внимание то, что максимальные частоты R1 наблюдаются в восточнославянских популяциях Европы (Semino et al., 2000a, Rosser et al., 2000). Наибольшее содержание западноевразийских линий Y хромосомы обнаружено в популяции долган (26%), наименьшее - у северных и вилюйских якутов (менее 5%). Гаплогруппы R1a и I встречаются в генофонде всех коренных этносов Якутии, тогда как гаплогруппы G, R1b, J, E3b выявлены в отдельных популяциях. Происхождение линий гаплогрупп I, G, E, типичных для Европы и Ближнего Востока, в генофонде коренных народов Якутии, скорее всего, связано с процессами метисации с русскими и, возможно, другими европейскими этносами, заселившими территорию Якутии начиная с XVII в. Линии гаплогруппы I, присутствующие во всех популяциях Якутии, с высокими частотами обнаружены практически во всех восточноевропейских популяциях, не обнаружены в Западной Сибири (за исключением популяций западных эвенков и ненцев) (Karafet et al., 2002; Wells et al., 2001), с низкой частотой встречаются в некоторых популяциях Южной Сибири (Derenko et al., 2006). Также очевидно недавнее проникновение линий гаплогрупп E3b и G в генофонд долган, самого молодого этноса Сибири, сложившегося в период летописной истории с XVII по XIX вв. Согласно данным переписей и исторических документов в состав долган вошли линии русских старожилов Сибири (Долгих, 1963). Гаплогруппа E3b не обнаружена в других популяциях Якутии, Южной и Западной Сибири (за исключением тувинцев), но присутствует в небольших количествах во всех восточноевропейских популяциях. Повышенная частота гаплогруппы G характерна для целого ряда популяций Кавказа (Nasidze et al., 2004) и Ближнего Востока (Semino et al., 2000; Cinnioglu et al., 2004), из восточноевропейских популяций встречается с небольшими частотами у русских и украинцев.
Что касается гаплогрупп R1 а и J, выявленных в изученных популяциях, то не исключается более древнее происхождение части этих линий из южных регионов Алтае-Саянского нагорья: эти гаплогруппы широко распространены в популяциях Южной Сибири и Монголии (Karafet et al., 2002; Katoh et al., 2004; Derenko et al., 2006).
3.3. Разнообразие микросателлитных гаплотипов Y хромосомы в популяциях Якутии. Компаундные гаплотипы Y хромосомы были определены по шести микросателлитным STR-маркерам (DYS19, DYS388, DYS390, DYS391, DYS392, DYS393) у 317 индивидов. Медианные сети N3 (N1C)- и С3-гаплотипов изученных популяций представлены на рис.9.
.jpg)
Звездообразная структура филогенетической сети с высокой частотой центрального гаплотипа 14-12-23-11-16-14 во всех популяциях якутов указывает на ярко выраженный эффект основателя, характерный для этноса в целом, что полностью согласуется с ранее опубликованными данными (Zerjal et al., 1997; Пузырев, 2003; Pakendorf et al., 2002, 2006). Время коалесценции N3 (N1C) гаплотипов, рассчитанное для центральных, вилюйских и северных якутов, а также в целом для всех трех популяций равно 1600±1070, 1600±820, 3210±1150 и 1960±950 лет, соответственно. Спектр N3 (N1C) линий более разнообразен у северных якутов в сравнении с двумя другими этногеографическими группами и соответственно выше значение времени коалесценции. Более высокий уровень разнообразия линий может возникать в результате более длительного эволюционного развития в изолированных условиях, но в данном случае, на наш взгляд, наиболее вероятным объяснением этому может быть смешанное происхождение северных якутов, вобравших N3 (N1C - линии соседних этносов -эвенков, эвенов и юкагиров. Из 16 выявленных N3 (N1C)-гаплотипов с линиями якутов пересекаются 5 линий эвенков, эвенов и юкагиров. При сравнении этих линий с N3 (N1C)-гаплотипами различных популяций (Rootsi et al., 2006) было установлено, что две из них филогенетически более близки к гаплотипам чукчей и эскимосов. Если исключить эти гаплотипы как, вероятно, имеющие другое происхождение, из расчетов, то значения времени коалесценции понижаются: для вилюйских якутов 1160±430 лет, для северных якутов 2150±680 лет и для всех трех популяций якутов 1540±580 лет.
В изученной нами выборке только у 2-х эвенков обнаружены N3 (N1C)-гаплотипы, отличные от линий якутов. Вероятно, происхождение большинства N3 (N1C -линий в мужском генофонде эвенков РС(Я) может объясняться смешением с якутами. Косвенным подтверждением этому служит то, что в других этногеографических группах эвенков обнаружены низкие частоты N3 (N1C)-гаплогруппы: в китайской популяции - 0 (Karafet et al., 2002), западной - от 0 (Pakendorf et al., 2006; Karafet et al., 2002) до 16% (Деренко, 2006).
C3c-гаплотипы представлены более широким спектром и имеют достоверно более высокие частоты в генофондах тунгусоязычных этносов Якутии - эвенков и эвенов.
Более высокий уровень разнообразия STR-линий наблюдается в популяциях эвенков, эвенов и юкагиров. Максимальные значения показателя генетического разнообразия линий выявлены в популяции юкагиров. У якутов наблюдается наиболее низкий уровень генетического разнообразия линий Y-хромосомы как по частотам гаплогрупп, так и по разнообразию STR-линий.
3.4. Филогеография мажорных гаплогрупп. Гаплогруппа N имеет широкое распространение в Евразии от Северной Европы до Дальнего Востока с максимальными частотами в северных широтах (Zerjal et al., 1997; Karafet et al., 2002; Tambets et al., 2004; Rootsi et al., 2007). Высокая частота гаплогруппы N является отличительной особенностью популяций Чукотки, Якутии, Западной Сибири, а также народов финно-угорской языковой группы, составляя примерно 3090% от общего пула Y хромосом в различных популяциях. Гаплогруппа N имеет три ветви - N1, N2 (N1B) и N3 (N1C), определяющиеся мутациями в локусах М128, Р43 и ТАТ, соответственно. Парагруппа N* встречается с низкими частотами, главным образом, в популяциях Юго-Восточной Азии - на Фиджи, Борнео, Камбодже, Южном Китае, Японии. Ареал распространения N* линий достигает Южной Сибири. В Индии не выявлено линий этой парагруппы (Karafet et al., 2002 ; Hammer et al., 2006; Sengupta et al., 2006; Степанов, 2006; Rootsi et al., 2007). N1-линии обнаружены с низкими частотами в некоторых популяциях Средней Азии, в Северном Китае и Корее. Эта ветвь гаплогруппы N является самой редкой и недостаточно изученной к настоящему времени (Karafet et al., 2002; Hammer et al., 2006; Rootsi et al., 2007).
Гаплогруппа N2 (N1B) имеет неравномерный паттерн распределения в популяциях Сибири и Европы. Наибольшие частоты гаплогруппы N2 (N1B) наблюдаются в популяциях Западной Сибири - 92% у нганасан, 78% у энцев и 74% у тундровых ненцев (Karafet et al., 2002). В Европе частота гаплогруппы N2 (N1B) составляет более 20% в некоторых популяциях Волго-Уральского региона (удмурты, коми) (Tambets et al., 2004). Западной границей ареала N2 (N1B) линий является Финляндия, где эта гаплогруппа встречается со следовыми частотами ~ 0.4% (Lappalainen et al., 2006; Rootsi et al., 2007).
Из ветвей гаплогруппы N наиболее распространенной является ветвь N3 (N1C), которая выявляется почти во всех популяциях Северной Евразии. В южных широтах - в Китае, Корее, Японии N3 (N1C -линии встречаются крайне редко. Хотя частота гаплогруппы N3 (N1C) в популяциях Северного Китая низкая, показатель разнообразия STR-линий (0.26) (Sengupta et al., 2006) выше, чем на Алтае и Волго-Уральском регионе (0.16 и 0.17, соответственно) (Rootsi et al., 2007), что указывает на то, что регионом, откуда произошла экспансия N3 (N1C -носителей, является Северный Китай, а не Южная Сибирь. Время генерации разнообразия N3 (N1C)-линий в Северном Китае оценено в 11800 ± 6800 лет (Rootsi et al., 2007), среднее значение попадает на границу между плейстоценом и голоценом, но большие значения стандартной ошибки требуют осторожной интерпретации полученных данных. Филогеография N* и NO* линий и присутствие N* линий главным образом в Юго-Восточной Азии указывает на то, что этот регион является наиболее вероятной прародиной гаплогруппы N, откуда линии этой гаплогруппы распространились по территории Евразии. Гаплогруппа N имеет недавнее происхождение. Значения времени генерации микросателлитного разнообразия N* линий также указывают на период позднего плейстоцена - раннего голоцена (от 11900 ± 2500 до 12600 ± 3100 лет). Для сравнения - время коалесценции N-линий, включая ветви N1, N2 (N1B), N3 (N1C), рассчитанное по вариабельности 15 STR локусов, несколько выше - 19400 ± 4800 лет.
Если исключить из расчета хромосомы, которые произошли от одного основателя (N3 (N1C -хромосомы якутов и N2 (N1B)-E европейские хромосомы), то время генерации микросателлитного разнообразия равно 14200 ± 4000 лет. Гаплогруппа N не выявлена в популяциях коренных народов Америки. Этот факт указывает на то, что в генофонде предковых сибирских популяций, заселивших американский континент по последним оценкам 12-17 тыс. лет назад (Zegura et al., 2004) гаплогруппа N отсутствовала, хотя не исключается возможность того, что N-линии могли исчезнуть в результате эффекта генетического дрейфа.

На рис.10 представлена медианная сеть, построенная по вариабельности 17 STR-локусов в N-O линиях различных популяций. В гаплогруппе N3 (N1C) выделяются два подкластера: в первый входят преимущественно популяции Волго-Уральского региона, а также отдельные линии тюркоязычных популяций (тувинцы и алтайцы), во второй - популяции балтов (эстонцы, карелы и вепсы), восточных славян (русские, украинцы) и словаков, чукчей и эскимосов. N3 (N1C) линии якутов образуют отдельную специфическую ветвь второго подкластера.
STR-линии гаплогруппы N2 (N1B) также образуют два отдельных подкластера, образованные азиатскими и европейскими гаплотипами, названные, соответственно, N2 (N1B)-A и N2 (N1B)-E. Можно предположить, что подкластер N2(N1B)-A, включающий линии сибирских популяций, имеет более раннее происхождение, а европейские N2(N1B)-E хромосомы образовались из одного гаплотипа-основателя.
Гаплогруппа С имеет неравномерное распространение по территории Евразии в направлении с юга на север, от Австралии до Сибири. Эта гаплогруппа отсутствует в Африке и Европе, западной границей ее распространения являются Средняя Азия и Ближний Восток (Karafet et al., 1999). С низкими частотами гаплогруппа С встречается также в популяциях татар и русских (Степанов, 2002; Balanovsky et al., 2008), что, по-видимому, связано с миграциями на восток степных кочевников Монголии, Южной Сибири и Средней Азии в относительно недавнее время (скифо-сибирское, последующее гунно-сарматское, древнетюркское время и в эпоху Золотой Орды). В гаплогруппе С выделяют три ветви - С1, С2 и С3, определяемые маркерами М8, М38 и М217, соответственно. Ветвь С1 идентифицирована в работе Т.Карафет и соавторов как гаплотип №19, не встречающийся в популяциях Юго-Восточной и Центральной Азии (Karafet et al., 2001). Кластер С2 распространен в популяциях Индонезии и Новой Гвинеи (Kayser et al., 2003). Основная ветвь гаплогруппы С - С3 - имеет наибольшие частоты в Южной Сибири и Монголии, Дальнем Востоке и Якутии. Кластер С3 является вторым по частоте в сибирских популяциях после гаплогруппы N. В отличие от N-линий C3-линии больше представлены в южных широтах Сибири. Предковые типы этой гаплогруппы возникли на территории Восточной Азии, после чего распространились в южном (С2) и северном (С3) направлениях. Часть С3-линий проникла в Америку, где они составляют около 6% (Zegura et al., 2004) от общего количества линий Y хромосомы.
Ветвь С3 включает линии С3а, С3Ь и С3с. С3b-линии, определяемые дополнительной мутацией в локусе Р39, характерны для популяций коренных жителей Америки. Линии С3b (С-Р39) находятся на третьем месте по частоте в аборигенных популяциях Америки после Q-M3 и Q-P36 линий (Zegura et al., 2004; Карафет, 2006). Линии С3с (С-М48), как уже упоминалось выше, характерны для тунгусо- и монголоязычных популяций Сибири, казахов, коряков и ительменов Камчатки. Время коалесценции линий гаплогруппы С, рассчитанное М.Хаммером и СЗегура (Hammer and Zegura, 2002), равно 27500±10100 лет. По данным В. Н.Харькова (2005) возраст генерации микросателлитного разнообразия гаплогруппы С3 имеет значение 26380±4530 лет, ветви С3с - 12080±3760 лет. Время коалесценции С3-линий в Якутии 14600 ± 5100 лет, ветви С3с - 8600 ± 5600.
3.5. Генетические взаимоотношения между популяциями Якутии по полиморфизму Y хромосомы.

На рис.11 А показано положение изученных популяций в пространстве двух главных компонент, описывающих 80% вариабельности линий Y хромосомы. Так же как и в случае мтДНК, все популяции якутов образуют отдельный компактный кластер вследствие высокой частоты гаплогруппы N3 (N1C). Популяция северных якутов несколько удалена от двух других групп из-за повышенного содержания гаплогруппы С (до 14%). Второй кластер образован популяциями долган, эвенков и эвенов, генофонд которых характеризуется высоким содержанием С3с-линий. Популяция юкагиров расположена отдельно из-за вклада специфических линий C3*(C3c), Q, I1b (I2a), O и отсутствия гаплогрупп N2 (N1B) и E. Характер кластеризации популяций в пространстве двух главных компонент по частотам гаплогрупп Y хромосомы сходен с таковым по частотам гаплогрупп мтДНК. Исключение составляет популяция эвенов, митохондриальный генофонд которых более близок к генофонду юкагиров, тогда как состав отцовских линий сходен с эвенкийским.
Высокое количество совпадающих STR-линий наблюдается между популяциями якутов, эвенков и эвенов (60-91%), якутов и долган (50-79%). Наибольшая вероятность совпадения линий выявлена между популяциями центральных и вилюйских якутов. Между популяциями юкагиров и долган не выявлено совпадающих STR-линий. Значения вероятности совпадения линий минимальны между популяцией юкагиров и всеми другими изученными популяциями, таким образом, спектр STR-линий юкагиров характеризуется наибольшими отличиями от других популяций Якутии. Наибольшая вероятность совпадения линий у юкагиров с эвенами.Дендрограмма, построенная на основе матрицы генетических расстояний, показывает отдаленность юкагиров и эвенов от популяций якутов (рис.11Б).
Результаты оценки генетических взаимоотношений между популяциями, как и в случае мтДНК, обнаруживают наиболее тесное генетическое родство между центральными и вилюйскими якутами. Максимальные различия выявляются между популяцией юкагиров, с одной стороны, и популяциями центральных и вилюйских якутов, с другой.
3.6. Анализ молекулярных различий между популяциями РС(Я) (AMOVA)
Уровень генетической подразделенности изученных популяций по вариабельности типов Y хромосомы намного выше, чем по линиям мтДНК. Если рассматривать все 7 популяций как одну группу, то на межпопуляционные различия приходится 20.7% вариабельности гаплогрупп Y хромосомы (табл.5).
Высокое количество совпадающих STR-линий наблюдается между популяциями якутов, эвенков и эвенов (60-91%), якутов и долган (50-79%). Наибольшая вероятность совпадения линий выявлена между популяциями центральных и вилюйских якутов. Между популяциями юкагиров и долган не выявлено совпадающих STR-линий. Значения вероятности совпадения линий минимальны между популяцией юкагиров и всеми другими изученными популяциями, таким образом, спектр STR-линий юкагиров характеризуется наибольшими отличиями от других популяций Якутии. Наибольшая вероятность совпадения линий у юкагиров с эвенами.Дендрограмма, построенная на основе матрицы генетических расстояний, показывает отдаленность юкагиров и эвенов от популяций якутов (рис.11Б).
Результаты оценки генетических взаимоотношений между популяциями, как и в случае мтДНК, обнаруживают наиболее тесное генетическое родство между центральными и вилюйскими якутами. Максимальные различия выявляются между популяцией юкагиров, с одной стороны, и популяциями центральных и вилюйских якутов, с другой.
3.6. Анализ молекулярных различий между популяциями РС(Я) (AMOVA)
Уровень генетической подразделенности изученных популяций по вариабельности типов Y хромосомы намного выше, чем по линиям мтДНК. Если рассматривать все 7 популяций как одну группу, то на межпопуляционные различия приходится 20.7% вариабельности гаплогрупп Y хромосомы (табл.5).

Высокий уровень межпопуляционных различий определяется, в первую очередь, особенностями мужского генофонда популяций якутов, для которых характерно крайне низкое разнообразие типов Y хромосомы, связанное с эффектом основателя по мужской линии. Оценка подразделенности мужского генофонда популяций Якутии по STR-локусам представляется более реальной (9.8%). Уровень генетической дифференциации популяций Якутии по микросателлитным маркерам ниже, чем в Средней Азии (13.46%), Западной Сибири (11.58%), но выше, чем в Восточной Европе (0.77%) (Степанов, 2002).
Степень генетической дифференциации популяций РС(Я) была определена также с помощью метода, предложенного P.W.Hedrick (2005). Значение стандартизированной Gst, рассчитанной по частотам гаплогрупп Y-хромосомы почти в 4 раза выше, чем по мтДНК - 45.9% и 12.3%, таким образом, различная степень генетической вариабельности этих систем не может быть объяснением значительных отличий в степени генетической дифференциации мужского и женского генофонда популяций Якутии.Степень генетической поразделенности популяций Якутии по разнообразию типов Y хромосомы (20.7%) ниже, чем в Западной Сибири (42.2%), сопоставима с такими регионами как Дальний Восток и Северный Китай (26.7%), Южная Сибирь (19.8%), Чукотка и Камчатка (22.1%) (табл.5).
3.7. Генетические взаимоотношения между популяциями Якутии и популяциями соседних регионов по Y хромосоме оценивали с помощью метода главных компонент и филогенетического анализа. В пространстве двух главных компонент, описывающих 37.6% вариабельности частот гаплогрупп Y-хромосомы, рассматриваемые популяции размещаются в 2 кластерах (рис.12).  Результаты кластерного анализа демонстрируют близость популяций Якутии к популяциям Южной Сибири, Дальнего Востока, Чукотки и Камчатки, отражая сходное высокое содержание гаплогрупп N3 (N1C) и С в их генофонде. |
4. Палеолитическое население Якутии - сохранились ли его гены?
По представлениям археологов с древнейших времен на территории Якутии происходила последовательная смена множества культур в результате волн миграций населения из более южных областей. Большая часть территории Якутии не была покрыта ледниковыми полями и была доступна для заселения и во время максимума сартанского оледенения 18-20 тыс. лет назад. Любопытным представляется вопрос, сохранились ли в генофонде современных этносов Якутии следы древних миграций эпохи верхнего палеолита, которые привели к заселению Берингии и, позднее, Америки? Результаты анализа мтДНК указывают на то, что гаплогруппы, характерные для популяций Америки (А2, В2, C1, D1, X2a, D4h3 и С4с), не обнаруживаются в митохондриальном генофонде популяций Якутии. Гаплогруппа D2, которая связана с более поздними миграциями в Берингию (Tamm et al., 2007), найдена в Якутии с низкой частотой (1.2%) и представлена единственной D2b-линией, распространенной в азиатских популяциях.
Что касается пула линий Y хромосомы, следует отметить, что он характеризуется крайне низким уровнем разнообразия, учитывая тот факт, что популяция якутов с ярко выраженным эффектом основателя составляет подавляющую часть коренного населения Якутии. Гаплогруппы Q и C3*, характерные для аборигенных популяций Америки (Seielstad et al. 2003; Zegura et al. 2004), вероятно, должны иметь более древний «возраст» в сибирских популяциях, т. к. они должны были присутствовать на этой территории еще во времена колонизации Америки, более 14 тыс. лет назад (Карафет, 2006). Более высокая частота этих гаплогрупп наблюдается в популяции юкагиров (27% C3* и 9% Q),. Так как содержание этих типов более древнего происхождения в генофонде популяций Якутии низкое (менее 7%), то можно предположить, что очень незначительная часть палеолитических линий древнего населения Якутии сохранилась в генофонде современного населения.
5. Анализ полиморфизма аутосомных ALU-инсерций в популяциях
якутов и эвенков
Если по полиморфизму митохондриальной ДНК и Y хромосомы можно получить характеристики женского и мужского генного пула, то исследование полиморфизма аутосомных Alu-инсерций позволяет определить некоторые особенности разнообразия генома человека в популяциях в целом. Мы проанализировали распределение частот и параметры генетического разнообразия 8 аутосомных Alu-инсерций (ACE, ApoA1, PV92, TPA25, NBC27, NBC102, NBC148, NBC182) в популяциях якутов и эвенков РС(Я) в сравнении с популяциями Волго-Уральского региона (башкиры, татары, коми, марийцы, мордва, удмурты), Средней Азии (казахи, узбеки и уйгуры) и Северного Кавказа (карачаевцы, кумыки, кубанские ногайцы и караногайцы) (Хуснутдинова, 2006; Kutuev et al., 2006). Частоты Alu-инсерций представлены в табл.6.

Действительно, частоты изученных Alu-инсерций имеют выраженный градиент: для локусов ACE, PV92, TPA25, NBC102, NBC148 в направлении с востока на запад в сторону снижения частот, а для локусов ApoA1, NBC27 и NBC182 - в сторону повышения. Данные по частотам Alu-инсерций в популяциях Якутии укладываются в закономерности распределения, выявленные ранее по другим регионам. Полученные результаты согласуются с литературными данными о высокой частоте инсерций в локусах PV92 и NBC148 в азиатских популяциях (Watkins et al., 2001, 2003; Хусаинова, 2004; Хуснутдинова, 2006). Из изученных полиморфизмов наиболее информативными маркерами дифференцированности западно- и восточно-евразийских популяций являются ACE, NBC102 и NBC27. В Якутии, которая является самым восточным регионом из рассмотренных, наблюдаются наиболее высокие средние частоты локусов ACE (0.607), PV92 (0.663), NBC102 (0.595), NBC148 (0.334) и наименьшие - по локусам NBC27 (0.191) и NBC182 (0.468).
Уровень гетерозиготности значительно варьирует в различных популяциях: минимальное значение по всем 8 локусам характерно для популяций мордвы (0.364) и марийцев (0.375) Волго-Уральского региона, максимальное - для уйгуров (0.464) и казахов (0.457) Средней Азии. Для популяций Якутии выявлены промежуточные значения - 0.422 для якутов и 0.427 для эвенков. Наибольший уровень генетического разнообразия в популяциях Якутии выявлен по локусам TPA25 и NBC182 (~0.5).
Для популяций якутов и эвенков установлены наиболее низкие значения Gst=0.001, что указывает на близкое генетическое родство между ними. Для сравнения - для популяций Волго- Уральского региона Gst=0.034, Северного Кавказа Gst=0.014, Средней Азии Gst=0.002. Cтепень подразделенности между популяциями Якутии и Средней Азии Gst=0.007.
Анализ потока генов в различных популяциях Азии, проведенный на основе полиморфизма тех же 8 Alu-инсерций, свидетельствует о низком генном потоке в популяциях Якутии, что согласуется с полученными ранее результатами для якутов Усть-Алданского улуса (Степанов, 2002; Хитринская, 2003).
6. Анализ ДНК-локусов, сцепленных с наследственными заболеваниями впопуляциях РС(Я)
6.1. Полиморфизм (СТС)п-локуса гена миотонинпротеинкиназы в популяциях РС(Я) и Средней Азии. Изучение полиморфизма (CTG)n-участка 3’-области гена миотонинпротеинкиназы (DMPK) представляет значительный интерес как в популяционном, так и в медицинском аспекте для исследования механизмов экспансии. Увеличение числа (CTG)n триплетов приводит к развитию миотонической дистрофии (МД) - наследственного нейромышечного заболевания. У здоровых людей число CTG-повторов варьирует от 5 до 37, тогда как на мутантных хромосомах их число может достигать нескольких тысяч (Brook et al., 1992). В Республиканском генетическом регистре среди наследственных болезней нервной системы миотоническая дистрофия занимает второе место по частоте после спиноцеребеллярной атаксии 1 типа. Высокое накопление МД наблюдается только в популяции якутов (Коротов, 1994; Сухомясова, 2005), тогда как в популяциях других коренных этносов Якутии (юкагиров, эвенов, эвенков, долганов) и у пришлого населения это заболевание практически не встречается.
Особенности аллельного спектра (CTG)n -локуса гена DMPK в популяциях якутов. Спектр аллельных вариантов (CTG)n-локуса гена DMPK был определен в популяциях Якутии и Средней Азии. Из исследованных популяций необходимо выделить якутов в силу особенностей распределения частот аллелей (CTG)n повторов, которое может быть охарактеризовано как одномодальное - пик в 11-14 повторов составляет 88-89%, тогда как этот локус имеет, как правило, тримодальное распределение в большинстве популяций мира. Значения показателя ожидаемой гетерозиготности (hexp) свидетельствуют о низком уровне генетического разнообразия в популяциях якутов по сравнению с другими популяциями Евразии, что характерно для изолированных этносов и островных популяций, происходящих от небольшого количества прародителей.
Частота аллеля (CTG)5, широко распространенного в европейских популяциях, у якутов составляет от 0.030 до 0.059 в различных этногеографических группах. Эти данные хорошо согласуются с результатами наших исследований спектра гаплотипов митохондриальной ДНК и Y хромосомы, согласно которым содержание западноевразийских линий в генофонде якутов достаточно низкое Содержание нестабильных аллелей с числом повторов более 19 у якутов составляет от 4 до 5.3% в различных этногеографических группах, в популяциях мира это значение варьирует от 0 до 15.1%. Частота аллелей с нестабильными повторами сравнительно невысокая, поэтому нет оснований предполагать, что существующие в популяции мутации гена DMPK имеют разное происхождение и возникли из нормальных аллелей большой длины. Накопление МД в якутской популяции, по видимому, обуславливается эффектом основателя и не связано с аллелями, имеющими число триплетных повторов >19.
Анализ генетических взаимоотношений между популяциями Якутии и Средней Азии по полиморфизму (СТО) повторов гена DMPK. Для оценки генетических взаимоотношений исследованных популяций по полиморфизму (CTG)n повтора гена DMPK использовали метод главных компонент и филогенетический анализ.


Проведен филогенетический анализ взаимоотношений популяций Якутии и Средней Азии (рис.13Б). На дендрограмме дифференцируются две ветви, одну из которых составляют популяции Якутии, а вторую - популяции Средней Азии. В кластере популяций РС(Я) наиболее близки к популяциям Средней Азии оказались эвенки, затем последовательно ответвляются три популяции якутов - вилюйские якуты, затем якуты центральной и, наконец, северной групп улусов. К популяции северных якутов примыкает популяция юкагиров. Последний, наиболее удаленный от корня кластер образуют долганы и эвены.
С помощью обоих подходов показаны существенные различия между популяциями двух регионов - Якутии и Средней Азии, для первых характерна большая выраженность показателей, характерных для монголоидов Азии, тогда как для вторых - близость к европейским популяциям. Как факторный, так и кластерный анализ выявляют тесное генетическое родство между центральными и вилюйскими якутами и обособленность популяции северных якутов от двух других этногеографических групп. По данным филогенетического анализа, северные якуты входят в один кластер с малочисленными народами Севера - юкагирами, эвенами и долганами. Исследованная в данной работе популяция эвенков дистанцирована от других популяций Якутии и проявляет наибольшее сходство с южными азиатскими популяциями. Степень генетической дифференциации по (CTG)n-локусу у популяций Якутии выше
(Fst=0.020), чем у популяций Средней Азии (Fst=0.008). Более высокий уровень генетической подразделенности населения Якутии может объясняться большей территориальной изолированностью популяций коренных этносов РС(Я), изначально малочисленных в сравнении с народами Средней Азии.
6.2. Скрининг мутаций C282Y и H63D в гене гемохроматоза в популяции якутов. Гемохроматоз - наследственное заболевание с нарушением метаболизма железа, характеризующееся прогрессирующим накоплением этого элемента в паренхиматозных органах, что приводит к развитию цирроза печени, диабета, кардиопатий, артрозов и эндокринных нарушений. Заболевание развивается к 30-40 годам, частота встречаемости составляет 1 на 300-500 человек, а носителей - 1 на 810 человек (до 10%) в европейских популяциях (Balan et al., 1994; Edwards et al., 1988; Adams et al., 1995; Baer et al., 1995).
Гемохроматоз имеет высокую генетическую гетерогенность. Основная форма гемохроматоза - тип 1 - связана с мутациями гена HFE, локализованного на 6 хромосоме (Feber et al., 1996; Simon et al., 1980). В настоящее время в гене HFE выявлены более 10 мутаций, сцепленных с заболеванием (Human Gene Mutation Database). Две миссенс-мутации являются мажорными: замена цистеина на тирозин в 282 положении HFE белка (C282Y), что нарушает образование дисульфидных мостиков, необходимых для стабилизации структуры белка; вторая мутация - замена гистидина на аспарагиновую кислоту в положении 63 аминокислотной цепи (H63D). Идентификация мутаций в гене HFE обеспечивает раннюю диагностику заболевания, популяционный скрининг носительства и проведение медико-генетического консультирования. Ранняя диагностика и лечение гемохроматоза предотвращает повреждение тканей и уменьшает смертность среди пациентов.
Возраст мутации C282Y оценен в 2000 лет (Rochette et al., 1999). Существуют гипотезы о североевропейском происхождении этой мутации, распространение которой связывают с кельтским населением (Byrnes et al., 2001; Rochette et al., 1999) или викингами (Milman and Pedersen, 2003). В популяции якутов мутация C282Y не обнаружена, что согласуется с данными о распространении этой мутации с запада на восток (Edwards et al., 1988; Adams et al., 1995; Burke et al, 1998; Тимковская, 2006; Хусаинова, 2006) и достаточно низким содержанием западноевразийского компонента по линиям мтДНК и Y хромосомы (менее 10%) в генофонде якутов.
Частота гетерозиготного носительства мутации H63D в популяции якутов достаточно высокая - 23.8%. Известно, что роль мутации H63D в развитии гемохроматоза меньше, чем C282Y мутации (UK Haemochromatosis Consortium, 1997; Goldwurm et al., 1997). Исследования показывают, что эта мутация ассоциирована с гемохроматозом, но со слабой пенетрантностью (Beutler et al., 1996; Beutler, 1997; Sham et al., 1997; Camaschella et al., 1997; Burk et al., 1998). Есть данные, что у гомозигот по мутации H63D развивается легкая форма гемохроматоза с более поздним проявлением. Мутация H63D наиболее часто встречается в странах Средиземноморья, на Ближнем Востоке, реже встречается в Азии и крайне редко - в популяциях Африки, Центральной и Южной Америки (Rochette et al., 1999; Merryweather-Clark et al., 2000; Тимковская, 2006; Хусаинова, 2006).
Учитывая обнаруженную частоту встречаемости мутаций С282Y и H63D в выборке здоровых индивидов из популяции якутов можно предположить, что частота гемохроматоза в Якутии должна быть намного ниже, чем в европейских регионах.
6.3. Структура генофонда якутской популяции и распространенность наследственных болезней: причинно-следственные взаимосвязи. Исходя из выявленных особенностей структуры генофонда якутов, можно предполагать некоторые особенности в распространении наследственных болезней в популяции.
1. Небольшое содержание западноевразийских линий в генофонде якутов (менее 10%) предполагает низкую распространенность в популяции наследственных болезней, имеющих редкую частоту возникновения и специфичных для популяций Европы. Действительно, по данным анализа спектра наследственных болезней в РС(Я) (Ноговицына, 1999; Тарская, 2004), в популяции якутов практически отсутствуют такие заболевания, как фенилкетонурия и муковисцидоз, широко распространенные в Европе. С другой стороны, любопытным является факт распространения болезни Фридрейха в популяции якутов. Это заболевание с аутосомно-рецессивным типом наследования выявлено в 7 якутских семьях (Тарская, 2004; Максимова, 2006a), частота встречаемости 1:36331. В азиатских популяциях это заболевание встречается крайне редко (Pandolfo, 2003; Chattopadhyay, 2004). Происхождение болезни Фридрейха в якутской популяции может быть связано как с процессами недавней метисации с пришлым русскоязычным населением, так и иметь более древние южносибирские корни.
2. Небольшая численность предковой популяции якутов с последующей значительной экспансией позволяет предположить, что для наследственных болезней, имеющих редкую частоту возникновения и получивших распространение в популяции, как правило, должен наблюдаться эффект родоначальника или присутствие в генах, ответственных за наследственные болезни, лишь одной мажорной мутации. В этой связи любопытно рассмотреть результаты некоторых исследований по эпидемиологии и происхождению таких наследственных нейродегенеративных болезней, характерных для популяции якутов, как спиноцеребеллярная атаксия 1 типа (QJ/d), миотоническая дистрофия (МД), окулофарингеальная дистрофия (ОФМД), а также редкого наследственного заболевания - синдрома 3-М. Частоты этих заболеваний в якутской популяции достигают максимальных значений в сравнении с другими популяциями мира (СЦА1 - 1:2590 (Платонов, 2003), МД - 1:4690 (Сухомясова, 2005), ОФМД - 1:16010 (Максимова, 2007), 3-М-синдрома - 1:8775 (Maksimova et al., 2007)). Ранее установлено, что происхождение СЦА1 типа в якутской популяции связано с эффектом родоначальника, при этом время распространения мутированного гена в Якутии оценивается исследователями в 915-1110 лет (Осаковский, Шатунов, 2004) и 1500-1750 лет (Гурьев, 2004). Результаты наших исследований нормального полиморфизма гена миотонической дистрофии в популяциях РС(Я) косвенно свидетельствуют о том, что накопление МД в якутской популяции так же обуславливается эффектом основателя (Федорова, 2005). Возможность эффекта основателя в распространении ОФМД и 3-М-синдрома в якутской популяции высказывалась в исследованиях Н.Р.Максимовой и соавт. (Максимова, 2007; Maksimova et al., 2007).
Характерной особенностью митохондриального генофонда коренных народов Якутии является выраженное преобладание гаплогрупп C и D, широко распространенных в популяциях Северной Азии.
2. Основную долю в генофонде популяций Якутии по Y-хромосоме составляют восточноевразийские гаплогруппы N, C и гаплогруппа R1, характерная для популяций Европы, Южной Сибири и Средней Азии. Содержание гаплогрупп J, I, F, E3b, O, Q, G в пуле Y-хромосом составляет в сумме 5%.
3. Западноевразийские гаплогруппы представлены в популяциях Якутии ограниченным спектром линий: их суммарное содержание варьирует от 0 до 14% по данным мтДНК, и от 3 до 26% по данным Y хромосомы. Сравнительный анализ с линиями русских и других европейских популяций указывает на то, что часть этих линий имеет более древнее палеоевропеоидное происхождение, не связанное с недавней, в масштабах эволюционного времени, миграцией русскоязычного населения в Восточную Сибирь, начиная с XVII в.
4. По составу линий мтДНК и Y-хромосомы установлено тесное генетическое родство между центральными и вилюйскими якутами, большая генетическая близость популяций якутов к эвенкам, отдаленность якутов от юкагиров;
5. Популяции якутов характеризуются предельно низким уровнем разнообразия линий Y-хромосомы (71-93% мужчин в разных этногеографических группах являются потомками одного N3 (N1C-основателя), относительно высоким разнообразием линий мтДНК, высокой частотой гаплогруппы D5a2, низким уровнем генетических различий между популяциями по мтДНК и Y-хромосоме. Характерными особенностями популяций эвенков (Средней Сибири, Китая, Якутии) является высокая частота гаплогруппы C3c мужского генофонда, незначительное количество совпадающих линий мтДНК между популяциями, высокий уровень генетической дифференциации по обеим маркерным системам. Популяции юкагиров отличаются относительно высоким содержанием гаплогрупп Q и C3* Y-хромосомы, характерным для популяций Америки и древнего населения Сибири эпохи верхнего палеолита. Результаты анализа спектра линий эвенов и долган подтверждают смешанное происхождение этих этносов. Митохондриальный генофонд эвенов более близок к генофонду юкагиров, тогда как состав линий Y-хромосомы сходен с эвенкийским. По составу гаплотипов мтДНК и Y-хромосомы долганы ближе к эвенкам, чем к якутам, мужской генофонд долган отличается высоким содержанием западноевразийских линий (26%).
6. Особенности генетических портретов популяций Якутии по линиям Y хромосомы более ярко выражены, чем по мтДНК, что определяется в основном эффектом основателя в популяциях якутов и эффектом патрилокальности. Выявлен низкий уровень генетических различий между популяциями Якутии в сравнении с другими регионами Сибири по данным полиморфизма мтДНК. Степень генетической подразделенности между популяциями Якутии по разнообразию гаплогрупп Y хромосомы ниже, чем в Западной Сибири и сопоставима с такими регионами, как Дальний Восток, Южная Сибирь, Чукотка и Камчатка.
7. Филогеография мажорных гаплогрупп мтДНК и Y хромосомы свидетельствует о заселении территории Якутии из Южной Сибири, Монголии и Северного Китая.
По спектру линий мтДНК популяции Якутии наиболее близки к популяциям Южной Сибири, дистанцированы от палеоазиатских популяций Чукотки и Камчатки. По вариабельности частот гаплогрупп Y-хромосомы популяции Якутии группируются вместе с популяциями Южной Сибири, Чукотки и Камчатки, удалены от популяций Западной Сибири.
8. Анализ древней мтДНК из якутских погребений XVIII в. указывает на преемственность этих линий в популяции якутов за последние 300 лет. Результаты филогенетического анализа мтДНК двух человек из погребения эпохи позднего неолита (4100+100 л.н.-3300±100 л.н.) не соответствуют ранее выдвинутым гипотезам о генетической связи поздненеолитического населения Якутии с современными палеоазиатскими этносами.
9. Анализ полиморфизма 8 аутосомных Alu-инсерций в популяциях якутов и эвенков свидетельствует о незначительной степени дифференциации между ними, относительной близости к популяциям Средней Азии и низком уровне потока генов извне по сравнению с популяциями других регионов. Cтепень дифференциации между популяциями якутов и эвенков по Alu-локусам намного ниже, чем по линиям Y хромосомы и более сопоставима с уровнем подразделенности по линиям мтДНК, что объясняется большей эффективной численностью и меньшей генетической вариабельностью Alu-локусов в сравнении с системами Y хромосомы и мтДНК.
10. Небольшое содержание европеоидного компонента в генофонде якутов (менее 10%) предполагает низкую распространенность в популяции наследственных болезней, имеющих редкую частоту возникновения и специфичных для популяций Европы. Малочисленность предковой популяции якутов с последующей значительной экспансией позволяют предположить, что для всех наследственных болезней, имеющих редкую частоту возникновения и, получивших распространение в популяции, как правило, должен наблюдаться эффект основателя или присутствие в генах, ответственных за наследственные болезни, одной мажорной мутации.
6.2. Скрининг мутаций C282Y и H63D в гене гемохроматоза в популяции якутов. Гемохроматоз - наследственное заболевание с нарушением метаболизма железа, характеризующееся прогрессирующим накоплением этого элемента в паренхиматозных органах, что приводит к развитию цирроза печени, диабета, кардиопатий, артрозов и эндокринных нарушений. Заболевание развивается к 30-40 годам, частота встречаемости составляет 1 на 300-500 человек, а носителей - 1 на 810 человек (до 10%) в европейских популяциях (Balan et al., 1994; Edwards et al., 1988; Adams et al., 1995; Baer et al., 1995).
Гемохроматоз имеет высокую генетическую гетерогенность. Основная форма гемохроматоза - тип 1 - связана с мутациями гена HFE, локализованного на 6 хромосоме (Feber et al., 1996; Simon et al., 1980). В настоящее время в гене HFE выявлены более 10 мутаций, сцепленных с заболеванием (Human Gene Mutation Database). Две миссенс-мутации являются мажорными: замена цистеина на тирозин в 282 положении HFE белка (C282Y), что нарушает образование дисульфидных мостиков, необходимых для стабилизации структуры белка; вторая мутация - замена гистидина на аспарагиновую кислоту в положении 63 аминокислотной цепи (H63D). Идентификация мутаций в гене HFE обеспечивает раннюю диагностику заболевания, популяционный скрининг носительства и проведение медико-генетического консультирования. Ранняя диагностика и лечение гемохроматоза предотвращает повреждение тканей и уменьшает смертность среди пациентов.
Возраст мутации C282Y оценен в 2000 лет (Rochette et al., 1999). Существуют гипотезы о североевропейском происхождении этой мутации, распространение которой связывают с кельтским населением (Byrnes et al., 2001; Rochette et al., 1999) или викингами (Milman and Pedersen, 2003). В популяции якутов мутация C282Y не обнаружена, что согласуется с данными о распространении этой мутации с запада на восток (Edwards et al., 1988; Adams et al., 1995; Burke et al, 1998; Тимковская, 2006; Хусаинова, 2006) и достаточно низким содержанием западноевразийского компонента по линиям мтДНК и Y хромосомы (менее 10%) в генофонде якутов.
Частота гетерозиготного носительства мутации H63D в популяции якутов достаточно высокая - 23.8%. Известно, что роль мутации H63D в развитии гемохроматоза меньше, чем C282Y мутации (UK Haemochromatosis Consortium, 1997; Goldwurm et al., 1997). Исследования показывают, что эта мутация ассоциирована с гемохроматозом, но со слабой пенетрантностью (Beutler et al., 1996; Beutler, 1997; Sham et al., 1997; Camaschella et al., 1997; Burk et al., 1998). Есть данные, что у гомозигот по мутации H63D развивается легкая форма гемохроматоза с более поздним проявлением. Мутация H63D наиболее часто встречается в странах Средиземноморья, на Ближнем Востоке, реже встречается в Азии и крайне редко - в популяциях Африки, Центральной и Южной Америки (Rochette et al., 1999; Merryweather-Clark et al., 2000; Тимковская, 2006; Хусаинова, 2006).
Учитывая обнаруженную частоту встречаемости мутаций С282Y и H63D в выборке здоровых индивидов из популяции якутов можно предположить, что частота гемохроматоза в Якутии должна быть намного ниже, чем в европейских регионах.
6.3. Структура генофонда якутской популяции и распространенность наследственных болезней: причинно-следственные взаимосвязи. Исходя из выявленных особенностей структуры генофонда якутов, можно предполагать некоторые особенности в распространении наследственных болезней в популяции.
1. Небольшое содержание западноевразийских линий в генофонде якутов (менее 10%) предполагает низкую распространенность в популяции наследственных болезней, имеющих редкую частоту возникновения и специфичных для популяций Европы. Действительно, по данным анализа спектра наследственных болезней в РС(Я) (Ноговицына, 1999; Тарская, 2004), в популяции якутов практически отсутствуют такие заболевания, как фенилкетонурия и муковисцидоз, широко распространенные в Европе. С другой стороны, любопытным является факт распространения болезни Фридрейха в популяции якутов. Это заболевание с аутосомно-рецессивным типом наследования выявлено в 7 якутских семьях (Тарская, 2004; Максимова, 2006a), частота встречаемости 1:36331. В азиатских популяциях это заболевание встречается крайне редко (Pandolfo, 2003; Chattopadhyay, 2004). Происхождение болезни Фридрейха в якутской популяции может быть связано как с процессами недавней метисации с пришлым русскоязычным населением, так и иметь более древние южносибирские корни.
2. Небольшая численность предковой популяции якутов с последующей значительной экспансией позволяет предположить, что для наследственных болезней, имеющих редкую частоту возникновения и получивших распространение в популяции, как правило, должен наблюдаться эффект родоначальника или присутствие в генах, ответственных за наследственные болезни, лишь одной мажорной мутации. В этой связи любопытно рассмотреть результаты некоторых исследований по эпидемиологии и происхождению таких наследственных нейродегенеративных болезней, характерных для популяции якутов, как спиноцеребеллярная атаксия 1 типа (QJ/d), миотоническая дистрофия (МД), окулофарингеальная дистрофия (ОФМД), а также редкого наследственного заболевания - синдрома 3-М. Частоты этих заболеваний в якутской популяции достигают максимальных значений в сравнении с другими популяциями мира (СЦА1 - 1:2590 (Платонов, 2003), МД - 1:4690 (Сухомясова, 2005), ОФМД - 1:16010 (Максимова, 2007), 3-М-синдрома - 1:8775 (Maksimova et al., 2007)). Ранее установлено, что происхождение СЦА1 типа в якутской популяции связано с эффектом родоначальника, при этом время распространения мутированного гена в Якутии оценивается исследователями в 915-1110 лет (Осаковский, Шатунов, 2004) и 1500-1750 лет (Гурьев, 2004). Результаты наших исследований нормального полиморфизма гена миотонической дистрофии в популяциях РС(Я) косвенно свидетельствуют о том, что накопление МД в якутской популяции так же обуславливается эффектом основателя (Федорова, 2005). Возможность эффекта основателя в распространении ОФМД и 3-М-синдрома в якутской популяции высказывалась в исследованиях Н.Р.Максимовой и соавт. (Максимова, 2007; Maksimova et al., 2007).
ВЫВОДЫ
1. На основании анализа полиморфизма ГВС1 и 40 диагностических участков кодирующей области мтДНК получена детальная характеристика структуры митохондриального генофонда популяций Якутии (якутов, эвенков, эвенов, юкагиров и долган). Подавляющее большинство митохондриальных линий (91%) в популяциях Якутии относится к восточноевразийским кластерам A, B, C, D, F, G, M7, M13а, Y, Z, 9% принадлежат к гаплогруппам H, U, HV1, J, T, W, имеющим широкое распространение в Европе и на Ближнем Востоке. Характерной особенностью митохондриального генофонда коренных народов Якутии является выраженное преобладание гаплогрупп C и D, широко распространенных в популяциях Северной Азии.
2. Основную долю в генофонде популяций Якутии по Y-хромосоме составляют восточноевразийские гаплогруппы N, C и гаплогруппа R1, характерная для популяций Европы, Южной Сибири и Средней Азии. Содержание гаплогрупп J, I, F, E3b, O, Q, G в пуле Y-хромосом составляет в сумме 5%.
3. Западноевразийские гаплогруппы представлены в популяциях Якутии ограниченным спектром линий: их суммарное содержание варьирует от 0 до 14% по данным мтДНК, и от 3 до 26% по данным Y хромосомы. Сравнительный анализ с линиями русских и других европейских популяций указывает на то, что часть этих линий имеет более древнее палеоевропеоидное происхождение, не связанное с недавней, в масштабах эволюционного времени, миграцией русскоязычного населения в Восточную Сибирь, начиная с XVII в.
4. По составу линий мтДНК и Y-хромосомы установлено тесное генетическое родство между центральными и вилюйскими якутами, большая генетическая близость популяций якутов к эвенкам, отдаленность якутов от юкагиров;
5. Популяции якутов характеризуются предельно низким уровнем разнообразия линий Y-хромосомы (71-93% мужчин в разных этногеографических группах являются потомками одного N3 (N1C-основателя), относительно высоким разнообразием линий мтДНК, высокой частотой гаплогруппы D5a2, низким уровнем генетических различий между популяциями по мтДНК и Y-хромосоме. Характерными особенностями популяций эвенков (Средней Сибири, Китая, Якутии) является высокая частота гаплогруппы C3c мужского генофонда, незначительное количество совпадающих линий мтДНК между популяциями, высокий уровень генетической дифференциации по обеим маркерным системам. Популяции юкагиров отличаются относительно высоким содержанием гаплогрупп Q и C3* Y-хромосомы, характерным для популяций Америки и древнего населения Сибири эпохи верхнего палеолита. Результаты анализа спектра линий эвенов и долган подтверждают смешанное происхождение этих этносов. Митохондриальный генофонд эвенов более близок к генофонду юкагиров, тогда как состав линий Y-хромосомы сходен с эвенкийским. По составу гаплотипов мтДНК и Y-хромосомы долганы ближе к эвенкам, чем к якутам, мужской генофонд долган отличается высоким содержанием западноевразийских линий (26%).
6. Особенности генетических портретов популяций Якутии по линиям Y хромосомы более ярко выражены, чем по мтДНК, что определяется в основном эффектом основателя в популяциях якутов и эффектом патрилокальности. Выявлен низкий уровень генетических различий между популяциями Якутии в сравнении с другими регионами Сибири по данным полиморфизма мтДНК. Степень генетической подразделенности между популяциями Якутии по разнообразию гаплогрупп Y хромосомы ниже, чем в Западной Сибири и сопоставима с такими регионами, как Дальний Восток, Южная Сибирь, Чукотка и Камчатка.
7. Филогеография мажорных гаплогрупп мтДНК и Y хромосомы свидетельствует о заселении территории Якутии из Южной Сибири, Монголии и Северного Китая.
По спектру линий мтДНК популяции Якутии наиболее близки к популяциям Южной Сибири, дистанцированы от палеоазиатских популяций Чукотки и Камчатки. По вариабельности частот гаплогрупп Y-хромосомы популяции Якутии группируются вместе с популяциями Южной Сибири, Чукотки и Камчатки, удалены от популяций Западной Сибири.
8. Анализ древней мтДНК из якутских погребений XVIII в. указывает на преемственность этих линий в популяции якутов за последние 300 лет. Результаты филогенетического анализа мтДНК двух человек из погребения эпохи позднего неолита (4100+100 л.н.-3300±100 л.н.) не соответствуют ранее выдвинутым гипотезам о генетической связи поздненеолитического населения Якутии с современными палеоазиатскими этносами.
9. Анализ полиморфизма 8 аутосомных Alu-инсерций в популяциях якутов и эвенков свидетельствует о незначительной степени дифференциации между ними, относительной близости к популяциям Средней Азии и низком уровне потока генов извне по сравнению с популяциями других регионов. Cтепень дифференциации между популяциями якутов и эвенков по Alu-локусам намного ниже, чем по линиям Y хромосомы и более сопоставима с уровнем подразделенности по линиям мтДНК, что объясняется большей эффективной численностью и меньшей генетической вариабельностью Alu-локусов в сравнении с системами Y хромосомы и мтДНК.
10. Небольшое содержание европеоидного компонента в генофонде якутов (менее 10%) предполагает низкую распространенность в популяции наследственных болезней, имеющих редкую частоту возникновения и специфичных для популяций Европы. Малочисленность предковой популяции якутов с последующей значительной экспансией позволяют предположить, что для всех наследственных болезней, имеющих редкую частоту возникновения и, получивших распространение в популяции, как правило, должен наблюдаться эффект основателя или присутствие в генах, ответственных за наследственные болезни, одной мажорной мутации.
СПИСОК
ОСНОВНЫХ ПУБЛИКАЦИЙ ПО ТЕМЕ ДИССЕРТАЦИИ
1. Fedorova S., Fatkhlislamova R., Khusnutdinova E. The analysis of polymorphism of CTG trinucleotide repeats in the gene of myotonic dystrophy in population of Yakuts // Human Genom Meeting. Shanghai. China. 2002. P.77-78.
2. Fatkhlislamova R., Fedorova S., Berdina L., Khusnutdinova E. Mutation analysis of the HFE gene in Eurasian populations in Russia // Human Genom Meeting. Shanghai. China. 2002. P.150.
3. Bermisheva M., Damba L., Golubenko M., Fedorova S., Zhadanov S., Tambets K., Kivisild T., Voevoda M., Stepanov V.A., Osipova L., Khusnutdinova E.K., Villems R. Turkic languages and the genes of turkic-speaking people - no general correlation in maternal lineages // Book of abstracts. Collegium Antropologicum. 13th congress of the European anthropological association. Zagreb. Croatia. 2002. V.26. P.21.
4. Khusnutdinova E., Bermisheva M, Malyarchuk B., Derenko M., Osipova L., Fedorova S., Villems R. Towards a comprehensive understanding of the East European maternal heritage in its phylogeographic context // Meeting Human Origin and Diseases.
USA. Cold Spring Harbor. 2002. P.87-88.
5. Федорова С.А., Бермишева М.А., Виллемс Р., Максимова Н.Р., Кононова С.К., Степанова С.К., Куличкин С.С., Хуснутдинова Э.К. Структура генофонда якутов по данным о полиморфизме митохондриальной ДНК // Якутский медицинский журнал. 2003. №1. С.16-21.
6. Fedorova S.A., Fatkhlislamova R.I., Sukhomiasova A.L., Maksimova N.R., Nikolaeva I.A., Khusnutdinova E.K. Study of CTG-trinucleotide repeats polymorphism in the myotonic dystrophy gene in Yakut and Evenk populations // Genosconference. Italy. 2003. P.10.
7. Федорова С.А., Бермишева М.А., Виллемс Р., Максимова Н.Р., Хуснутдинова Э.К. Анализ линий митохондриальной ДНК в популяции якутов // Молекулярная биология. 2003. Т.37. №4. С.643-653.
8. Хусаинова Р.И., Федорова С.А., Хуснутдинова Э.К. Скрининг Cys282Tyr и His63Asp мутаций в гене HFE у народов Волго-Уральского региона и Республики Саха (Якутия) // Медицинская генетика. 2003. №5. C.207-212.
9. Федорова С.А., Кононова С.К., Ноговицына А.Н. ДНК-диагностика наследственных болезней в Якутии // Х Российско-Японский Международный симпозиум медицинского обмена. Якутск. 2003. C.516-517.
Ю.Федорова С.А., Бермишева М.А., Виллемс Р., Максимова Н.Р., Хуснутдинова Э.К. Генофонд якутов по данным полиморфизма митохондриальной ДНК // Х Российско-Японский Международный симпозиум медицинского обмена. Якутск. 2003. C.517-518.
11.Сухомясова А.Л., Федорова С.А., Коротов М.Н., Максимова Н.Р., Алексеева С.П., Сидорова О.Г., Николаева И.А., Кононова С.К., Степанова С.К., Фатхлисламова Р.И., Хуснутдинова Э.К., Ноговицына А.Н. Миотоническая дистрофия в Республике Саха (Якутия): популяционные особенности и подходы к ДНК-тестированию // Якутский медицинский журнал. 2003. №2. С.12-17.
12.Кононова С.К., Федорова С.А., Коротов М.Н., Сидорова О.Г., Платонов Ф.А. К вопросам профилактики спиноцеребеллярной атаксии 1 типа в Якутии // Якутский медицинский журнал. 2003. №1. С.13-16.
13.Reidla M., Kivisild T., Metspalu E., Kaldma K., Tambets K., Tolk H.-V., Parik J.,
Loogvali E.-L., Derenko M., Malyarchuk B., Bermisheva M., Zhadanov S., Pennarun E., Gubina M., Golubenko M., Damba L., Fedorova S., Gusar V., Grechanina E., Mikerezi I., Moisan J.P., Chaventre A., Khusnutdinova E., Osipova L., Stepanov V., Voevoda M., Achilli A., Rengo C., Rickards O., De Stefano G.F., Papiha S., Beckman L., Janicijevic B., Rudan P., Anagnou N, Michalodimitrakis E., Koziel S., Usanga E., Geberhiwot T., Herrnstadt C., Howell N., Torroni A., Villems R. Origin and diffusion of mtDNA haplogroup X // Am. J. Hum. Genet. 2003. V.73. P.1178-1190.
14.Khusnutdinova E., Kutuev I., Bermisheva M., Malyarshuk B., Derenko M., Osipova L., Gubina M., Stepanov V., Fedorova S., Tolk H.-V., Tambets K., Metspalu E., Kivisild T., Villems R. Phylogeography of Y-chromosomal and mitochondrial lineages and language affinities of Eurasian populations // Human Genom Meeting. 2004. Berlin. P.141.
15.Sukhomyasova A.L., Maximova N.R., Nikolaeva I.A., Stepanova S.K., Fedorova S.A., Nogovitsina A.N. Myotonic dystrophy in Yakutia // European Human Genetics Сonference. Munich, Germany. 2004. P.124.
16.Sukhomyasova A.L., Maximova N.R., Korotov M.N., Nikolaeva I.A., Sidorova O.G., Fedorova S.A., Nogovitsina A.N. Congenital myotonic dystrophy in Yakutia // 11th International Symposium of the Japan-Russia Medical Exchange. Niigata, Japan. 2004. P.262.
17. Fedorova S.A., Khusnutdinova E.K., Villems R. Diversity of mtDNA and Y-chromosome lineagues in populations of Republic Sakha (Yakutia) // Human Genom Meeting. Kioto. Japan. 2005. Р.59.
18. Федорова С.А., Хусаинова Р.И., Кутуев И.А., Сухомясова А.Л., Николаева И.А., Куличкин С.С., Ахметова В.Л., Салимова А.З., Святова Г.С., Березина Г.М., Платонов Ф.А., Хуснутдинова Э.К. Полиморфизм CTG-повторов гена миотонинпротеинкиназы в популяциях Республики Саха (Якутия) и Средней Азии // Молекулярная биология. 2005. Т.39 №3. С.385-393.
19. Федорова С.А., Сухомясова А.Л., Николаева И.А., Куличкин С.С., Платонов Ф. А., Хуснутдинова Э.К. Аллельный полиморфизм гена миотонинпротеинкиназы в популяциях Республики Саха (Якутия) // Наука и образование. №2 (38). 2005. С.59-65.
20. Кононова С.К., Федорова С.А., Степанова С.К., Хуснутдинова Э.К. Подходы к ДНК-диагностике моногенных заболеваний в практике медико-генетической консультации Якутии // Наука и образование. №2 (38). 2005. С.104-111.
21. Федорова С. А., Хуснутдинова Э. К., Виллемс Р. Генофонд коренных этносов Якутии: анализ материнских и отцовских линий // Материалы международной конференции «Генетические аспекты патологии человека. Проблемы сохранения генофонда коренных народов Севера» Якутск. 2005. С.102-103.
22. Федорова С.А., Озава Т., Степанов А.Д., Жирков Э.К., Алексеева Л.Л., Адояан М., Парик Ю., Виллемс Р. Анализ линий древней митохондриальной ДНК в Якутии // Материалы международной конференции «Генетические аспекты патологии человека. Проблемы сохранения генофонда коренных народов Севера». Якутск. 2005. С.104-105.
23. Fedorova S.A., Ozawa T., Stepanov A.D., Jirkov E.K., Alekseeva L.L., Adoyan M., Parik J., Villems R. Analysis of ancient mitochondrial DNA lineages in Yakutia // The American Society of Human Genetics. 55th Annual Meeting. USA. Uta. 2005. P.197.
24. Kutuev I., Khusainova R., Karunas A., Yunusbayev B., Fedorova S., Lebedev Y.,Hunsmann G., Khusnutdinova E. From east to west: patterns of genetic diversity of populations living in four Eurasian regions.// Human Heredity. 2006. V.61. P.1-9.
25. Кононова С.К., Федорова С.А., Степанова С.К., Сидорова С.Г., Хуснутдинова Э.К. Организационные, методические и этические проблемы ДНК-диагностики моногенных заболеваний в практике медико-генетической консультации Якутии // Медицинская генетика. 2006. Т.5. C.14-19.
26. Rootsi S, Zhivotovsky L.A., Baldovic M., Kayser M., Kutuev I.A., Khusainova R., Bermisheva M.A., Gubina M., Fedorova S., Ilumae A.M., Khusnutdinova E.K., Osipova L.P., Stoneking M., Ferak V., Parik J., Kivisild T., Underhill P., Villems R. A counter-clockwise northern route of the Y-chromosome haplogroup N from Southeast Asia towards Europe // Eur. J. Hum. Genet. 2007. V.15. P.204-211.
27. Барашков Н.А., Джемилева Л.У., Федорова С.А., Максимова Н.Р., Сухомясова А.Л., Гуринова Е.Е., Кононова С.К., Терютин Ф.М., Федотова Э.Е., Ноговицына А.Н., Хуснутдинова Э.К. Вклад мутаций 35delG, 167delT и 235delC гена коннексина 26 (GJB2) в возникновении наследственной несиндромальной сенсоневральной тугоухости в Республике Саха (Якутия) // Медицинская генетика. 2007. №9. С.26-30.
28. Fedorova S.A., Platonov F.A., Khusnutdinova E.K., Villems R. Peculiarities of gene pool structure of indigenous populations of Yakutia and origin of some hereditary neurodegenerative disorders spread in Yakuts // III International Meeting on problems of viliuisk encephalomyelitis and others neurodegenerative disorders in Yakutia. Yakutsk. 2006. P.18-19.
29.Sukhomyasova A.L., Nazarenko L.P., Maksimova N.R., Danilova A.L., Korotov M.N., Nikolaeva L.A., Gurinova E.E., Fedorova S.A., Nogovicina A.N. Myotonic dystrophy in Yakutia // III International Meeting on problems of viliuisk encephalomyelitis and others neurodegenerative disorders in Yakutia. Yakutsk. 2006. P.23.
30. Федорова С.А., Хуснутдинова Э.К. Структура генофонда якутов и вопросы молекулярной эпидемиологии наследственных болезней // Материалы межрегиональной научно-практической конференции «Молекулярно-клеточные аспекты патологии человека на Севере». Якутск. 2007. С.43-44.
31. Tamm E., Kivisild T., Reidla M., Metspalu M., Smith D.G., Mulligan C.J., Bravi C.M., Rickards O., Martinez-Labarga C., Khusnutdinova E.K., Fedorova S.A., Golubenko M.V., Stepanov V.A., Gubina M.A., Zhadanov S.I., Ossipova L.P., Damba L., Voevoda M.I., Dipierri J.E., Villems R., Malhi R.S. Beringian standstill and spread of Native American founders // PLoS ONE. 2007. Issue 9, e829.
32. Федорова С.А., Степанов А.Д., Жирков Э.К., Адоаян М., Парик Ю, Озава Т., Виллемс Р. Анализ линий древней митохондриальной ДНК в Якутии // Молекулярная биология. 2008. Т.42. №3. С.445-453.
33. Федорова С.А. Генетические портреты народов Республики Саха (Якутия): анализ линий митохондриальной ДНК и Y-хромосомы. Якутск: Изд-во ЯНЦ СО РАН. 2008. 235 с.
1. Fedorova S., Fatkhlislamova R., Khusnutdinova E. The analysis of polymorphism of CTG trinucleotide repeats in the gene of myotonic dystrophy in population of Yakuts // Human Genom Meeting. Shanghai. China. 2002. P.77-78.
2. Fatkhlislamova R., Fedorova S., Berdina L., Khusnutdinova E. Mutation analysis of the HFE gene in Eurasian populations in Russia // Human Genom Meeting. Shanghai. China. 2002. P.150.
3. Bermisheva M., Damba L., Golubenko M., Fedorova S., Zhadanov S., Tambets K., Kivisild T., Voevoda M., Stepanov V.A., Osipova L., Khusnutdinova E.K., Villems R. Turkic languages and the genes of turkic-speaking people - no general correlation in maternal lineages // Book of abstracts. Collegium Antropologicum. 13th congress of the European anthropological association. Zagreb. Croatia. 2002. V.26. P.21.
4. Khusnutdinova E., Bermisheva M, Malyarchuk B., Derenko M., Osipova L., Fedorova S., Villems R. Towards a comprehensive understanding of the East European maternal heritage in its phylogeographic context // Meeting Human Origin and Diseases.
USA. Cold Spring Harbor. 2002. P.87-88.
5. Федорова С.А., Бермишева М.А., Виллемс Р., Максимова Н.Р., Кононова С.К., Степанова С.К., Куличкин С.С., Хуснутдинова Э.К. Структура генофонда якутов по данным о полиморфизме митохондриальной ДНК // Якутский медицинский журнал. 2003. №1. С.16-21.
6. Fedorova S.A., Fatkhlislamova R.I., Sukhomiasova A.L., Maksimova N.R., Nikolaeva I.A., Khusnutdinova E.K. Study of CTG-trinucleotide repeats polymorphism in the myotonic dystrophy gene in Yakut and Evenk populations // Genosconference. Italy. 2003. P.10.
7. Федорова С.А., Бермишева М.А., Виллемс Р., Максимова Н.Р., Хуснутдинова Э.К. Анализ линий митохондриальной ДНК в популяции якутов // Молекулярная биология. 2003. Т.37. №4. С.643-653.
8. Хусаинова Р.И., Федорова С.А., Хуснутдинова Э.К. Скрининг Cys282Tyr и His63Asp мутаций в гене HFE у народов Волго-Уральского региона и Республики Саха (Якутия) // Медицинская генетика. 2003. №5. C.207-212.
9. Федорова С.А., Кононова С.К., Ноговицына А.Н. ДНК-диагностика наследственных болезней в Якутии // Х Российско-Японский Международный симпозиум медицинского обмена. Якутск. 2003. C.516-517.
Ю.Федорова С.А., Бермишева М.А., Виллемс Р., Максимова Н.Р., Хуснутдинова Э.К. Генофонд якутов по данным полиморфизма митохондриальной ДНК // Х Российско-Японский Международный симпозиум медицинского обмена. Якутск. 2003. C.517-518.
11.Сухомясова А.Л., Федорова С.А., Коротов М.Н., Максимова Н.Р., Алексеева С.П., Сидорова О.Г., Николаева И.А., Кононова С.К., Степанова С.К., Фатхлисламова Р.И., Хуснутдинова Э.К., Ноговицына А.Н. Миотоническая дистрофия в Республике Саха (Якутия): популяционные особенности и подходы к ДНК-тестированию // Якутский медицинский журнал. 2003. №2. С.12-17.
12.Кононова С.К., Федорова С.А., Коротов М.Н., Сидорова О.Г., Платонов Ф.А. К вопросам профилактики спиноцеребеллярной атаксии 1 типа в Якутии // Якутский медицинский журнал. 2003. №1. С.13-16.
13.Reidla M., Kivisild T., Metspalu E., Kaldma K., Tambets K., Tolk H.-V., Parik J.,
Loogvali E.-L., Derenko M., Malyarchuk B., Bermisheva M., Zhadanov S., Pennarun E., Gubina M., Golubenko M., Damba L., Fedorova S., Gusar V., Grechanina E., Mikerezi I., Moisan J.P., Chaventre A., Khusnutdinova E., Osipova L., Stepanov V., Voevoda M., Achilli A., Rengo C., Rickards O., De Stefano G.F., Papiha S., Beckman L., Janicijevic B., Rudan P., Anagnou N, Michalodimitrakis E., Koziel S., Usanga E., Geberhiwot T., Herrnstadt C., Howell N., Torroni A., Villems R. Origin and diffusion of mtDNA haplogroup X // Am. J. Hum. Genet. 2003. V.73. P.1178-1190.
14.Khusnutdinova E., Kutuev I., Bermisheva M., Malyarshuk B., Derenko M., Osipova L., Gubina M., Stepanov V., Fedorova S., Tolk H.-V., Tambets K., Metspalu E., Kivisild T., Villems R. Phylogeography of Y-chromosomal and mitochondrial lineages and language affinities of Eurasian populations // Human Genom Meeting. 2004. Berlin. P.141.
15.Sukhomyasova A.L., Maximova N.R., Nikolaeva I.A., Stepanova S.K., Fedorova S.A., Nogovitsina A.N. Myotonic dystrophy in Yakutia // European Human Genetics Сonference. Munich, Germany. 2004. P.124.
16.Sukhomyasova A.L., Maximova N.R., Korotov M.N., Nikolaeva I.A., Sidorova O.G., Fedorova S.A., Nogovitsina A.N. Congenital myotonic dystrophy in Yakutia // 11th International Symposium of the Japan-Russia Medical Exchange. Niigata, Japan. 2004. P.262.
17. Fedorova S.A., Khusnutdinova E.K., Villems R. Diversity of mtDNA and Y-chromosome lineagues in populations of Republic Sakha (Yakutia) // Human Genom Meeting. Kioto. Japan. 2005. Р.59.
18. Федорова С.А., Хусаинова Р.И., Кутуев И.А., Сухомясова А.Л., Николаева И.А., Куличкин С.С., Ахметова В.Л., Салимова А.З., Святова Г.С., Березина Г.М., Платонов Ф.А., Хуснутдинова Э.К. Полиморфизм CTG-повторов гена миотонинпротеинкиназы в популяциях Республики Саха (Якутия) и Средней Азии // Молекулярная биология. 2005. Т.39 №3. С.385-393.
19. Федорова С.А., Сухомясова А.Л., Николаева И.А., Куличкин С.С., Платонов Ф. А., Хуснутдинова Э.К. Аллельный полиморфизм гена миотонинпротеинкиназы в популяциях Республики Саха (Якутия) // Наука и образование. №2 (38). 2005. С.59-65.
20. Кононова С.К., Федорова С.А., Степанова С.К., Хуснутдинова Э.К. Подходы к ДНК-диагностике моногенных заболеваний в практике медико-генетической консультации Якутии // Наука и образование. №2 (38). 2005. С.104-111.
21. Федорова С. А., Хуснутдинова Э. К., Виллемс Р. Генофонд коренных этносов Якутии: анализ материнских и отцовских линий // Материалы международной конференции «Генетические аспекты патологии человека. Проблемы сохранения генофонда коренных народов Севера» Якутск. 2005. С.102-103.
22. Федорова С.А., Озава Т., Степанов А.Д., Жирков Э.К., Алексеева Л.Л., Адояан М., Парик Ю., Виллемс Р. Анализ линий древней митохондриальной ДНК в Якутии // Материалы международной конференции «Генетические аспекты патологии человека. Проблемы сохранения генофонда коренных народов Севера». Якутск. 2005. С.104-105.
23. Fedorova S.A., Ozawa T., Stepanov A.D., Jirkov E.K., Alekseeva L.L., Adoyan M., Parik J., Villems R. Analysis of ancient mitochondrial DNA lineages in Yakutia // The American Society of Human Genetics. 55th Annual Meeting. USA. Uta. 2005. P.197.
24. Kutuev I., Khusainova R., Karunas A., Yunusbayev B., Fedorova S., Lebedev Y.,Hunsmann G., Khusnutdinova E. From east to west: patterns of genetic diversity of populations living in four Eurasian regions.// Human Heredity. 2006. V.61. P.1-9.
25. Кононова С.К., Федорова С.А., Степанова С.К., Сидорова С.Г., Хуснутдинова Э.К. Организационные, методические и этические проблемы ДНК-диагностики моногенных заболеваний в практике медико-генетической консультации Якутии // Медицинская генетика. 2006. Т.5. C.14-19.
26. Rootsi S, Zhivotovsky L.A., Baldovic M., Kayser M., Kutuev I.A., Khusainova R., Bermisheva M.A., Gubina M., Fedorova S., Ilumae A.M., Khusnutdinova E.K., Osipova L.P., Stoneking M., Ferak V., Parik J., Kivisild T., Underhill P., Villems R. A counter-clockwise northern route of the Y-chromosome haplogroup N from Southeast Asia towards Europe // Eur. J. Hum. Genet. 2007. V.15. P.204-211.
27. Барашков Н.А., Джемилева Л.У., Федорова С.А., Максимова Н.Р., Сухомясова А.Л., Гуринова Е.Е., Кононова С.К., Терютин Ф.М., Федотова Э.Е., Ноговицына А.Н., Хуснутдинова Э.К. Вклад мутаций 35delG, 167delT и 235delC гена коннексина 26 (GJB2) в возникновении наследственной несиндромальной сенсоневральной тугоухости в Республике Саха (Якутия) // Медицинская генетика. 2007. №9. С.26-30.
28. Fedorova S.A., Platonov F.A., Khusnutdinova E.K., Villems R. Peculiarities of gene pool structure of indigenous populations of Yakutia and origin of some hereditary neurodegenerative disorders spread in Yakuts // III International Meeting on problems of viliuisk encephalomyelitis and others neurodegenerative disorders in Yakutia. Yakutsk. 2006. P.18-19.
29.Sukhomyasova A.L., Nazarenko L.P., Maksimova N.R., Danilova A.L., Korotov M.N., Nikolaeva L.A., Gurinova E.E., Fedorova S.A., Nogovicina A.N. Myotonic dystrophy in Yakutia // III International Meeting on problems of viliuisk encephalomyelitis and others neurodegenerative disorders in Yakutia. Yakutsk. 2006. P.23.
30. Федорова С.А., Хуснутдинова Э.К. Структура генофонда якутов и вопросы молекулярной эпидемиологии наследственных болезней // Материалы межрегиональной научно-практической конференции «Молекулярно-клеточные аспекты патологии человека на Севере». Якутск. 2007. С.43-44.
31. Tamm E., Kivisild T., Reidla M., Metspalu M., Smith D.G., Mulligan C.J., Bravi C.M., Rickards O., Martinez-Labarga C., Khusnutdinova E.K., Fedorova S.A., Golubenko M.V., Stepanov V.A., Gubina M.A., Zhadanov S.I., Ossipova L.P., Damba L., Voevoda M.I., Dipierri J.E., Villems R., Malhi R.S. Beringian standstill and spread of Native American founders // PLoS ONE. 2007. Issue 9, e829.
32. Федорова С.А., Степанов А.Д., Жирков Э.К., Адоаян М., Парик Ю, Озава Т., Виллемс Р. Анализ линий древней митохондриальной ДНК в Якутии // Молекулярная биология. 2008. Т.42. №3. С.445-453.
33. Федорова С.А. Генетические портреты народов Республики Саха (Якутия): анализ линий митохондриальной ДНК и Y-хромосомы. Якутск: Изд-во ЯНЦ СО РАН. 2008. 235 с.