Вопреки ранним, и фактически необоснованным утверждениям, что R1b является «западноевропейской» гаплогруппой, предки которой жили в Европе 30 тысяч лет назад, и определенно были кроманьонцами, на самом деле европейский вариант R1b является относительно молодой гаплогруппой (в основном R1b1b2/M269), предок которой пришел в Европу из Азии не более 4500-5000 лет назад. Азиатский вариант этой гаплогруппы (в основном R1b1b1/M73) настолько отличается по мутациям от европейского, что это показывает, что их общий предок жил - скорее всего, в Азии - примерно 16 тысяч лет назад. Общие предки наиболее популярных в Европе субкладов - R1b1b2/R1b1c/M269, R1b1b2a1/R1b1c9/U106/S21/M405, R1b1b2a2g/R1b1c10/U152/S28
- жили примерно в одно и то же время, 4200 - 4500 лет назад. Даже баски, которых (без особых обоснований в отношении гаплогруппы R1b) считали старейшими жителями Европы, имеют общего предка по гаплогруппе R1b не ранее 4000-4600 лет назад.
Гаплогруппа R1b привлекает особое внимание любителей и профессионалов ДНК-генеалогии. Причина проста - ее больше всего у тех и у других, по крайней мере по данным, доступным в настоящее время. Эта гаплогруппа доминирует в Западной и Центральной Европе и на Британских островах, и поэтому ее чаще всего тестируют у тех, кто способен оплатить коммерческое определение гаплотипа и гаплогруппы. По ряду причин такими и оказываются жители Западной Европы.
По данным на начало 2008 года в базе данных YSearch было 44093 гаплотипа, и из них 17171, то есть почти 40%, принадлежали гаплогруппе R1b c подгруппами. Это показывает желание ивозможности европейцев узнать свои корни. Для сравнения - только 31 человек гаплогруппы R1a1 из России и Украины, из нескольких десятков миллионов, проявили пока такое сочетание желания и возможности.
Как часто бывает в ДНК-генеалогии, фактически только зарождающейся области науки, гаплогруппа R1b, как самая популярная, стала быстро обрастать легендами и фантазиями. Часть из них были основаны на скоропалительных и непроверенных результатах, тем не менее опубликованных в серьезных научных журналах и книгах. Часть появились вообще без всякого обоснования.
Наиболее устойчивая легенда говорит о том, что гаплогруппу R1b имели кроманьонцы, что эта гаплогруппа была в Европе 30-35 тысяч лет назад, ее обладатели охотились бок о бок с неандертальцами, и ее же обладатели оставили пещерные рисунки на юге Европы, которые датируются вплоть до 32 тысяч лет назад. В качестве примера особо древних носителей гаплогруппы R1b часто приводят басков. Видимо, потому что у них древний язык, не относящийся к индоевропейской языковой группе.
Вот как, например, описывает гаплогруппу R1b один из пионеров ДНК- генеалогии, Спенсер Уэллс, в своей недавней книге Deep Ancestry (2006): «Примерно 30 тысяч лет назад один из потомков клана, который находился на пути в Европу, получил мутацию М343, которая и определила его в новую гаплогруппу R1b. Потомки этого человека - прямые потомки кроманьонцев, которые доминировали в освоении Европы, создали знаменитые наскальные рисунки в пещерах на юге Франции».
Свою роль в этих легендах сыграли некорректные методы расчета «вариации» мутаций в гаплотипах, использование неких «популяционных» скоростей мутаций, в которых весьма произвольно и на основании некритичных допущений резко отдалялись времена жизни «общих предков» выборок гаплотипов, делались ошибки в отнесении гаплотипов к гаплогруппам, или расчеты проводились вообще без таких отнесений.
В данную работу не входит детальное рассмотрение того, как именно возникали легенды про гаплогруппу R1b, и в чем именно были ошибки в расчетах. Хотя следует отметить по меньшей мере три источника таких ошибок:
Гаплогруппа R1b привлекает особое внимание любителей и профессионалов ДНК-генеалогии. Причина проста - ее больше всего у тех и у других, по крайней мере по данным, доступным в настоящее время. Эта гаплогруппа доминирует в Западной и Центральной Европе и на Британских островах, и поэтому ее чаще всего тестируют у тех, кто способен оплатить коммерческое определение гаплотипа и гаплогруппы. По ряду причин такими и оказываются жители Западной Европы.
По данным на начало 2008 года в базе данных YSearch было 44093 гаплотипа, и из них 17171, то есть почти 40%, принадлежали гаплогруппе R1b c подгруппами. Это показывает желание ивозможности европейцев узнать свои корни. Для сравнения - только 31 человек гаплогруппы R1a1 из России и Украины, из нескольких десятков миллионов, проявили пока такое сочетание желания и возможности.
Как часто бывает в ДНК-генеалогии, фактически только зарождающейся области науки, гаплогруппа R1b, как самая популярная, стала быстро обрастать легендами и фантазиями. Часть из них были основаны на скоропалительных и непроверенных результатах, тем не менее опубликованных в серьезных научных журналах и книгах. Часть появились вообще без всякого обоснования.
Наиболее устойчивая легенда говорит о том, что гаплогруппу R1b имели кроманьонцы, что эта гаплогруппа была в Европе 30-35 тысяч лет назад, ее обладатели охотились бок о бок с неандертальцами, и ее же обладатели оставили пещерные рисунки на юге Европы, которые датируются вплоть до 32 тысяч лет назад. В качестве примера особо древних носителей гаплогруппы R1b часто приводят басков. Видимо, потому что у них древний язык, не относящийся к индоевропейской языковой группе.
Вот как, например, описывает гаплогруппу R1b один из пионеров ДНК- генеалогии, Спенсер Уэллс, в своей недавней книге Deep Ancestry (2006): «Примерно 30 тысяч лет назад один из потомков клана, который находился на пути в Европу, получил мутацию М343, которая и определила его в новую гаплогруппу R1b. Потомки этого человека - прямые потомки кроманьонцев, которые доминировали в освоении Европы, создали знаменитые наскальные рисунки в пещерах на юге Франции».
Свою роль в этих легендах сыграли некорректные методы расчета «вариации» мутаций в гаплотипах, использование неких «популяционных» скоростей мутаций, в которых весьма произвольно и на основании некритичных допущений резко отдалялись времена жизни «общих предков» выборок гаплотипов, делались ошибки в отнесении гаплотипов к гаплогруппам, или расчеты проводились вообще без таких отнесений.
В данную работу не входит детальное рассмотрение того, как именно возникали легенды про гаплогруппу R1b, и в чем именно были ошибки в расчетах. Хотя следует отметить по меньшей мере три источника таких ошибок:
(а) использование неверных скоростей мутаций в гаплотипах, например, «коэффициента Животовского», который примерно в три раза отдаляет «время жизни общего предка» (Zhivotovsky et al, 2004; Sengupta et al, 2006; Karlsson et al, 2006; Derenko et al, 2007; Gay den et al, 2007; Martinez et al, 2007; Contu et al, 2008),
(б) смешивание гаплотипов из разных гаплогрупп, и
(в) использование коротких гаплотипов, иногда состоящих всего из трех-четырех маркеров.
Генезис гаплогруппы R1b
Рассмотрим, когда же на самом деле носители гаплогруппы R1b появились в Европе, откуда они там появились или могли появиться. Оборот «на самом деле» отражает, конечно, уровень современных представлений в ДНК-генеалогии. С появлением принципиально новых данных картина может измениться. Но это - нормальный путь научного познания.
В целом генезис гаплогруппы R1b (c некоторыми подгруппами) выглядит сейчас следующим образом:
Генезис гаплогруппы R1b
Рассмотрим, когда же на самом деле носители гаплогруппы R1b появились в Европе, откуда они там появились или могли появиться. Оборот «на самом деле» отражает, конечно, уровень современных представлений в ДНК-генеалогии. С появлением принципиально новых данных картина может измениться. Но это - нормальный путь научного познания.
В целом генезис гаплогруппы R1b (c некоторыми подгруппами) выглядит сейчас следующим образом:
«Адам»
→M168 → M89 → M9 → M207 → M173 → M343 → P25 → P297/M73 → M269 → (S116; S21/U106/M405; S28/U152)
В начале цепочки - «хромосомный Адам», который положил начало современному человечеству. К нему сходятся генеалогические линии всех людей на Земле. Каждый мужчина на Земле унаследовал его Y- хромосому и гаплотип (то есть определенную конфигурацию нуклеотидных последовательностей в определенных локусах Y- хромосомы, которые называют маркерами), и последующие поколения добавляли лишь мутации в эти маркеры. В итоге мы имеем то разнообразие гаплотипов и гаплогрупп, которое наблюдается у современных людей.
Первая мутация, М168, дала сводную гаплогруппу С-R в будущей гаплогруппе R1b. Гаплогруппы А и В остались в Африке. Мутация М89 привела к укороченной сводной гаплогруппе F-R. Дальнейшая мутация еще более укоротила сводную гаплогруппу, до K-R. Мутация М207 дала гаплогруппу R. Мутация М271 привела к гаплогруппе R1. Наконец, мутация М343 дала мутацию R1b. Как будет показано ниже, это произошло примерно 16 тысяч лет назад, в Азии, на участке пути в Европу, о чем первые носители гаплогруппы R1b, конечно, не знали. Пару тысяч лет спустя носители гаплогруппы R1b были в Армении. 5-6 тысяч лет назад они были, по-видимому, на Средне-Русской равнине, и, возможно, это они оставили археологические культуры боевых топоров, она же культура шнуровой керамики, она же культура одиночных могил. Не исключено, что и курганную культуру, хотя здесь необоснованных гипотез предостаточно в отношении и гаплогруппы R1a1. Но данных пока нет. |
Здесь приходится постоянно говорить «возможно», потому что археологи не знают, к какому ДНК-генеалогическому роду относились те (и почти все остальные) культуры, и таких определений пока не проводилось. Поэтому я здесь строю возможную модель, которую в дальнейшем нужно проверять.
4500-5000 лет назад носители R1b прибыли в Европу, примерно тогда же, когда члены рода R1a1 начали экспансию с Балкан во все стороны света - к Атлантике, к Скандинавии, на северо-восток и восток, на территории современных России, Украины, Белоруссии, Прибалтики, и на юг, на Апеннины, Пелопоннес, в Малую Азию.
Вот это - схема, которую я буду обосновывать в данной работе. Она пока представляется крайне неортодоксальной, и является по сути принципиальной ревизией легенд о R1b в Европе 30 тысяч лет назад. По моим данным, никаких R1b в Европе не было ранее 5-6 тысяч лет назад. Или, если они и были, то следов не оставили, во всяком случае в современном населении Европы.
Более того, недавно появились данные, принципиально подтверждающие, что баски гаплогруппы R1b, которых среди басков более 90%, появились в Испании и Франции никак не ранее нескольких тысяч лет назад. Оказалось, что более 90% их имеют ген лактозной толерантности (что позволяет им пить молоко без серьезного дискомфорта), который впервые появился в Азии менее 10 тысяч лет назад, и оттуда принесен в Европу. Об этом подробнее - ниже.
Субклады R1b
Но вернемся к схеме мутаций. Мы остановились на мутации М343, которая привела к образованию гаплогруппы R1b. И далее остановимся на подгруппах. Дело в том, что мутации, которые можно использовать для классификации новых гаплогрупп, происходят в нас довольно часто. В Y-хромосоме 50 миллионов нуклеотидов, и даже при скорости мутации одна на миллион в поколение в каждом поколении может появляться до 50 мутаций, которые могут быть квалифицированы как снипы (SNP = Single Nucleotide Polymorphism). Подобных мутаций в людях уже выявлено более трех миллионов (The International HapMap Consortium, 2007). Так что в принципе в каждой семье может быть выявлена мутация, которая присуща только этой семье, идущая, например, от прадедушки. Или пра-прадедушки. Это очень удобно для проведения так называемых «фамильных исследований», как серьезное подспорье в классической генеалогии. Иначе говоря, все прямые мужские потомки определенного предка, который жил, скажем, триста лет назад, несут его снип, и определенный тест это покажет. Такие линии называются «субкладами», и они нумеруются в виде продолжения индекса основной гаплогруппы.
Ясно, что для R1b таких субкладов предложено больше всего, а именно уже более тридцати. Их может быть и сотни, и тысячи, и, видимо, в будущем так и будет. Пока же создание новых субкладов - инициатива специалистов в ДНК-генеалогии, которые делают выбор мутаций и соответствующих субкладов на основании ряда причин, включая и личные предпочтения.
Так вот, следующей за R1b подгруппой, или субкладом, является R1b1, который определяется мутацией P25. Потом идет R1b1a, мутация M18. Затем идет Р297/M73, субклад R1b1b, который недавно переименован в R1b1b1. Это - в основном азиатский субклад, от Турции до Сибири. Далее - R1b1b2, мутация М269 (ранее, по предыдущей номенклатуре, был R1b1c). Это - европейский вариант R1b.
И затем идет целая галерея субкладов, нисходящих от мутации М269. Они раньше именовались R1b1c1, R1b1c2, R1b1c3, и далее до R1b1c11. Теперь они именуются в развитие индекса R1b1b2, например, R1b1b2a2c, R1b1b2a2d, R1b1b2a2e, R1b1b2a2g и так далее. Каждому поставлена в соответствие мутация с определенным индексом, часто многозначным (например, R1b1b2a2 - это rs34276300), и по параллельной системе мутации еще обозначают индексами S28, S116, а также индексами типа U106, U152, U198 и так далее, причем маркеры могут сопровождаться значками + или - . Система весьма усложнена, и вдобавок постоянно ревизуется, причем параллельно разными администраторами и разными тестирующими компаниями, которые предпочитают свои индексы.
Я вынужден это здесь описывать, поскольку несколько таких субкладов проанализировал на время жизни общего предка. А именно, следующие (идут синонимы по записи):
R1b1b2 = R1b1c = M269 = S3 = S10 = S13 = S17
R1b1b2a1 = R1b1c9 = M405 = S21 = U106+
R1b1b2a2g = R1b1c10 = S28 = U152+
Некоторые индексы дублируются, потому что номенклатура еще не устоялась, и продолжает идти путаница. В номенклатуре 2008 года (http://www.isogg.org/tree/IS0GG_HapgrpR08.html) отмечено, что субклад R1b1b2a1 (бывший R1b1b1c9) имеет маркер S21/U106. А проект http://www.familytreedna.com/public/U106, с маркером R-U106/S21+, представляет почти исключительно гаплотипы субклада R1b1b2g. В номенклатуре 2008 года субклада R1b1b2g вообще нет. И таких примеров немало.
Для нашей цели это, строго говоря, неважно. Мы рассмотрим времена общих предков гаплогруппы R1b и субкладов, как бы их не называли, и выясним, насколько это времена жизни общих предков отличаются друг от друга.
Сравнение времени жизни общих предков европейских R1al и R1b
Перед этим - расчет приблизительный. В базе данных YSearch имеются более 17 тысяч гаплотипов гаплогруппы R1b с подгруппами, в подавляющем большинстве европейского происхождения. Можно для начала провести расчет времени жизни общего предка для всех этих тысяч гаплотипов, понимая, что этот расчет будет в значительной степени условным. С другой стороны, наличие такого количества гаплотипов даст хорошую статистику, обычно недостижимую другими методами. Итак, в какой степени «условным» будет расчет? И насколько гаплотипы группы R1b окажутся «моложе» или «старше» гаплотипов группы R1a1?
Гаплогруппа R1a1 - для сравнения
В предыдущей статье, посвященной гаплогруппе R1a1 (Клёсов, 2008а), мы показали, что общий предок многих гаплотипов этой гаплогруппы жил в Европе 4400-4800 лет назад, для некоторых территорий от 4200 лет назад (Чехия, Словакия, Швеция), до 5000-5200 лет назад (Венгрия, Ирландия), для некоторых регионов раньше, чем 4200 лет назад. Но в среднем по Европе этот срок составляет примерно 4500 лет назад. А что даст массивный расчет по базе данных YSearch? Это даст некоторое представление, насколько целесообразны подобные массивные расчеты.
Итак, в базе данных есть 672 шести-маркерных гаплотипа R1a1, опять в основном европейского происхождения, с базовым (предковым) гаплотипом в системе записи FTDNA (пропущенные аллели заменены символами Х)
13-25-16-10-Х-Х-Х-12-Х-Х-11-Х
Все 672 гаплотипа имеют 1092 мутации от базового. Это дает в среднем 1092/672/6 = 0.271 мутацию на маркер, что - с поправкой на возвратные мутации - соответствует 204 поколениям до общего предка, или 5100 лет.
В той же базе данных есть 664 12-маркерных гаплотипов гаплогруппы R1a1, с базовым гаплотипом
13-25-16-10-11-14-12-12-10-13-11-30
Как видно, предковый гаплотип тот же самый, что и для 6-маркерных гаплотипов, но более полный, 12-маркерный. Все 664 гаплотипа содержат 2180 мутаций, что дает 0.274 мутации на маркер, и соответствует 179 поколениям до общего предка, то есть 4475 лет. Среднее количество мутаций на маркер здесь больше, чем в 6- маркерных гаплотипах, но средние скорости мутации в 6-маркерных гаплотипах меньше, чем в 12-маркерных. Поэтому и расстояние до общего предка оказалось несколько меньше.
Наконец, в базе данных есть 417 25-маркерных гаплотипов R1a1, и во всех имеется 3095 мутаций от базового гаплотипа
13-25-16-10-11-14-12-12-10-13-11-30-15-9-10-11-11-24-14-20-32-12-15-15-16
Это дает 0.297 мутаций на маркер, что соответствует 198 поколениям, или 4950 годам до общего предка. Интересно, что этот предковый 25- маркерный гаплотип в точности соответствует предковому гаплотипу русских (восточных) славян, хотя их было всего 26 гаплотипов из 417 , или всего 6%, во всей базе данных. Иначе говоря, славянский предковый гаплотип в точности равен общеевропейскому, что, впрочем, показано раньше (Клёсов, 2008a) на основе другого материала, не привлекая массированные расчеты, как делаю здесь.
Видно, что все эти величины укладываются в интервал времен жизни общих предков гаплогруппы R1a1, рассчитанных по разным европейским регионам (Клёсов, 2008a). То есть подобные массированные расчеты дают разумные величины времен жизни общего предка.
Гаплогруппа R1b
Применим тот же подход для гаплогруппы R1b (c подгруппами). Для 17171 шестимаркерных гаплотипов
13-24-14-11-Х-Х-Х-12-Х-Х-13-Х
имеется 21027 мутаций, что дает 0.204 мутации на маркер, и это соответствует 146 поколениям, или 3650 годам до общего предка всех 17 тысяч гаплотипов R1b в базе данных. Заметим, что общий предок R1b в Европе (а именно к Европе относится подавляющее количество гаплотипов R1b в базе данных) «моложе» общего предка R1a1. Это видно и по среднему количеству мутаций на маркер (0.271 для R1a1
Для 12-маркерных гаплотипов, которых в базе данных 16959, базовый гаплотип R1b следующий
13-24-14-11-11-14-12-12-12-13-13-29
Как видно, он опять согласуется с предковым 6-маркерным гаплотипом. Эти гаплотипы имеют суммарно 50040 мутаций, что дает 0.246 мутаций на маркер, и приводит к 158 поколениям, или 3950 годам до общего предка. Таким образом, и по 12-маркерным гаплотипам общий предок гаплогруппы R1b в Европе «моложе», чем общий предок гаплогруппы R1a1.
В базе данных имеется 12090 25-маркерных гаплотипов R1b, и их базовый (предковый) гаплотип, полученный минимизацией мутаций по каждому маркеру (Клёсов, 2008b)
13-24-14-11-11-14-12-12-12-13-13-29-17-9-10-11-11-25-15-19-29-15-15-17-17
позволил выявить 83 240 мутаций по всем гаплотипам. Это дает 0.275 мутаций на маркер, и приводит к 180 поколениям, или 4500 лет до общего предка.
Обратим внимание, что для гаплотипов R1a1 6-маркерные гаплотипы дали самый высокий интервал времени до общего предка по сравнению с 12- и 25-маркерными гаплотипами, а для гаплотипов R1b - самый низкий. Это понятно, поскольку 6-маркерные гаплотипы больше других подвержены колебаниям по числу мутаций. Я и привел их здесь в основном только для того, чтобы продемонстрировать этот результат. Тем не менее, даже 6-маркерные гаплотипы дают в целом разумные результаты по оценке времен жизни общего предка. На основании более точных 12- и 25-маркерных гаплотипов можно заключить, что общий предок европейских R1b жил примерно на 500 лет позже европейских же R1a1, а именно 4200±400 лет назад.
Как видно, это совсем не 30 тысяч лет назад, как говорят нам «научные» легенды. R1b - «младший брат» R1a1 в Европе.
Но это была, как я отмечал, примерная оценка возраста общего предка европейских R1b. Рассмотрим конкретные выборки гаплотипов более внимательно.
Европейские гаплотипы R1b двух основных генеалогических линий
На рис. 1 приведено дерево 37-маркерных европейских гаплотипов гаплогруппы R1b, в количестве 104. Они были выбраны неупорядоченным способом из всего массива R1b, и все были помечены в базе данных YSearch как "R1b (tested)".
Видно, что дерево разделяется на две части, в левой, более «молодой» части 75 гаплотипов, в правой части 24 гаплотипа, и еще малая ветвь справа из пяти гаплотипов (между 241 и 267) не обрабатывалась по причине малочисленности ветви.
При рассмотрении только первых 12 маркеров все дерево имеет один предковый гаплотип:
13-24-14-11-11-14-12-12-12-13-13-29
Это именно тот гаплотип, который приведен выше и который был получен при обработке почти 17 тысяч гаплотипов R1b. Это - самый распространенный европейский гаплотип, и он называется «Атлантический модальный гаплотип», или АМГ. Все 104 12- маркерных гаплотипа имеют 268 мутаций, что соответствует 135 поколений, или 3375 лет до общего предка.

При увеличении числа маркеров до 25 базовые гаплотипы начинают расходиться. Проявляются два общих предка, отстоящих по времени на 700 лет, и имевшие одинаковые 12-маркерные гаплотипы, но уже несколько разные 25-маркерные. В 25-маркерным формате их базовые гаплотипы выглядят следующим образом (мутационные различия обозначены жирным шрифтом): 13-24-14-11-11-14-12-12-12-13-13-29-17-9-9-11-11-25-15-19-29-15-15-16-17 13-24-14-11-11-14-12-12-12-13-13-29- 17-9-10-11-11-25-15-19-30-15-15-17-17 Приведенный выше 25-маркерный гаплотип, полученный из всего массива базы данных из тысяч гаплотипов - усредненный между ними. Верхний базовый гаплотип соответствует правой верхней части дерева на рис. 1, из 24 гаплотипов, нижний базовый гаплотип - левой части, из 75 гаплотипов. Правая часть дерева имеет 169 мутаций на 24 гаплотипа, что соответствует 185 поколениям, или 4625 годам до общего предка. Левая часть дерева - 460 мутаций на 75 гаплотипов, что соответствует 158 поколениям, или 3950 годам до общего предка. Как видно, эти данные близки к тем, которые были получены выше при массированном анализе более 10 тысяч гаплотипов базы данных. Как обычно и получается, результат мало зависит от величины выборки, когда выборка достигает даже небольших величин, зачастую 20-50 гаплотипов. Разница во времени между общими предками этих двух 25-маркерных гаплотипов составляет 700 лет. Это соответствует примерно 1.2 мутациям разницы, если один общий предок является прямым потомком другого. Для показанных выше двух гаплотипов эта разница составляет 0.90+0.15+0.06 = 1.11. Совпадение прекрасное. Более «молодые» гаплотипы (на рис. 1 слева) происходят от субклада «старшего». «Старший» - гаплотипы на рис. 1 справа - это продукт гаплогруппы R1b или R1b1, или R1b1b, или R1b1b2 (R1b1c). «Младший» субклад - это соответствующий нисходящий, как показано на схеме выше. Итак, самый древний общий предок R1b по этим данным прибыл в Европу примерно 4600 лет назад. В это же время род R1a1 уже осваивал новые территории, тоже по всей Европе, расходясь от своей балканской «прародины», где род R1a1 жил уже более семи тысяч лет. Так что это за молодой субклад, нисходящий от R1b, с общим предком, жившим в Европе примерно 4000 лет назад? Попытаемся его определить. Время жизни общих предков субкладов гаплогруппы R1b1b2
|

В этом собрании 284 полных 25-маркерных гаплотипа (и выше по числу маркеров), из них подавляющая часть гаплотипов с британскихостровов и из США, потомков жителей британских островов. Их в собрании три четверти всех гаплотипов. Остальные, примерно по десятку-полтора - из разных европейских регионов - Центральная Европа (9 гаплотипов), северо-восток Европы (12 гаплотипов), северозападная Европа (22 гаплотипа), Скандинавия (14 гаплотипов), юговосточная и юго-западная Европа (по три гаплотипа), и ряд гаплотипов без конкретного отнесения. Дерево 67-маркерных гаплотипов, построенное по этим данных, приведено на рис. 2.

Естественно, что дерево с меньшим разрешением, а именно дерево 25- маркерных гаплотипов, тоже указывает на одного общего предка (рис. 3).
По данным дерева на рис. 3 было рассчитано, что базовый (предковый) гаплотип субклада R1b-U106/S21 - следующий:
13-23-14-11-11-14-12-12-12-13-13-29- 17-9-10-11-11-25-15-19-29-15-15-17-17
Он всего лишь на одну мутацию (отмечено жирным шрифтом) отличается от обобщенного предкового гаплотипа, полученного выше при массивной обработке базы данных YSearch, на три мутации от предкового гаплотипа материнской гаплогруппы (рис. 1 справа), и на две мутации от предкового гаплотипа «молодого» субклада на рис. 1 (слева).
Все 284 25-маркерных гаплотипа содержат 1853 мутации от этого предкового гаплотипа, что соответствует 0.261 мутации на маркер, и транслируется в 4225 лет до общего предка всех 284 носителей этих гаплотипов.
Те же 12-маркерные гаплотипы содержат 729 мутаций от базового гаплотипа, что соответствует 0.214 мутациям на маркер, что дает 3375 лет до общего предка. Это - с поправкой на возвратные мутации. Без поправки «возраст» общего предка составил бы 729/284/0.022 = 117 поколений, или 2925 лет. Поправка увеличивает этот возраст на 450 лет, до 3375 лет.
900 лет - довольно большая разница в «возрасте» предков, полученным по 12- и 25-маркерным гаплотипам. Дополнительная проверка основывалась на числе идентичных гаплотипов во всей выборке, которые и соответствуют предковым (базовым) гаплотипам. Таких среди 284 гаплотипов оказалось 21. Общий предок жил ln(284/21)/0.022 = 118 поколений назад (без поправки на возвратные мутации), то есть практически идентично величине 117 поколений, полученной по мутациям. С поправкой это даст те же 3375 лет.
Действительно, расхождение результатов расчетов между 12- и 25- маркерными гаплотипами наблюдалось нами и ранее. Но часто расхождения нет, и данные практически идентичны для двух серий. Это означает, что проблема не в величинах используемых констант скоростей мутаций, но в особенностях конкретных серий гаплотипов - либо 12-, либо 25-маркерных. Эта проблема еще не решена в ДНК- генеалогии. Можно, конечно, начинать отбрасывать некоторые маркеры, но этим будет внесен произвол в расчеты. В данном случае проблемы особой нет, поскольку мы решаем принципиальный вопрос - когда R1b пришли в Европу - в каменном веке, 30 тысяч лет назад, или в бронзовом, 3-5 тысяч лет назад. Пока данные говорят о втором варианте, причем с большой степенью достоверности.
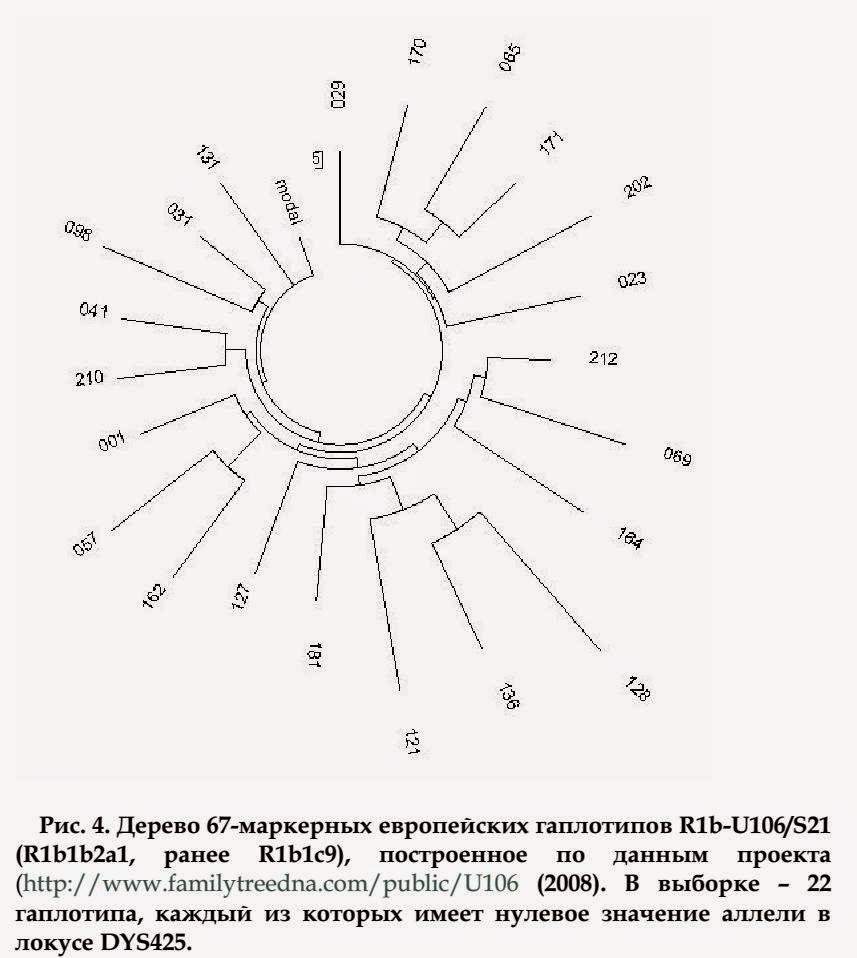
Субклад R1b1b2a1/U106/S21, с нулевым значением DYS425
Подборка гаплотипов субклада R1b-U106/S21 позволила проанализировать интересное явление. 22 гаплотипа из 306 имеют нулевое значение аллели в DYS425 (48-й по счету маркер в 67- маркерном гаплотипе). Это - результат так называемого эффекта RecLOH (Recombinational Loss of Heterozygosity), когда информация в локусе Y-хромосомы (в данном случае) при переписывании теряется. Поскольку это нулевое значение, раз приобретенное, передается по наследству, то интересно было узнать время жизни общего предка данных 22 человек, а значит - время, когда это обнуливание аллели произошло.
На рис. 4 показано дерево гаплотипов этих 22 человек. Дерево симметричное, и соответствует одному общему предку. Время жизни предка рассчитывалось по серии 25-маркерных гаплотипов носителей гаплотипов с нулевым значением. Предковый гаплотип их на первых 25 маркерах отличался на одну мутацию (выделено жирным шрифтом) от предкового гаплотипа для остальных объектов выборки:
13-23-14-11-11-14-12-12-12-13-13-29- 17-9-10-11-11-24-15-19-29-15-15-17-17
Иначе говоря, нулевая мутация в DYS425 пошла от человека, у которого уже случилась мутация в маркере DYS447 от обычной аллели 25 к 24.
Все 22 гаплотипа имели 54 мутации на 12-маркерных гаплотипах, и 118 мутаций на 25-маркерных гаплотипах. Это дает соответственно 128 и 135 поколений до общего предка, или 3200 и 3375 лет, соответственно.
Субклад R1b1b2a2g/U152/S28
Переходим еще к одному субкладу, R1b1c10/R1b1b2h/R1b1b2a2g (http://www.davidkfaux.org/R1b1c10_Data.htm), он же субклад S28/U152. Это - субклад, параллельный тому, что представлен на рис. 3. Они исходят из одного «материнского» субклада R1b1c, но на рис. 3 представлен с9, а на рис. 5 - с10.
Среди 184 гаплотипов субклада, представленных на рис. 5, есть восемь идентичных 12-маркерных гаплотипов. Согласно принципам ДНК- генеалогии, их рассматривают как базовые, или предковые гаплотипы, наиболее сохранившиеся, в которых со времени жизни общего предка ни произошло ни одной мутации. Они уже могут быть использованы для расчета времени до общего предка (Клёсов, 2008b) по формуле ln(184/8)/0.022 = 143 поколения (без поправки на возвратные мутации). В тех же 184 12-маркерных гаплотипов имеется 568 мутаций, что дает 568/184/0.022 = 140 поколений (также без поправки). То, что число поколений практически одинаково по двум совершенно разным способам расчета показывает, что данный субклад действительно имеет одного общего предка. Об этом же свидетельствует и вид дерева гаплотипов на рис. 5, которое не содержит выраженных ветвей.
С поправкой на возвратные мутации для 12-маркерных гаплотипов число поколения до общего предка отодвигается от 140-143 до 167 поколений, что соответствует 4175 годам. 184 25-маркерных гаплотипов содержат 1236 мутаций, что соответствует 175 поколениям, то есть 4375 лет до общего предка.
Как видно, в этом случае время жизни общего предка, рассчитанное по 12- и 25-маркерным гаплотипам почти одинаково, а именно 4175 и 4375 лет.
Из этого можно сделать вывод, что по 25-маркерным гаплотипам субклады R1b1c9 (U106) и R1b1c10 (U152) образовались практически в одно и то же время, а именно 4225 и 4375 лет назад, соответственно.

| 25-маркерные базовые (предковые) гаплотипы этих субкладов выглядят следующим образом: 13-23-14-11-11-14-12-12-12-13-13-29- 17-9-10-11-11-24-15-19-29-15-15-17-17 13-24-14-11-11-14-12-12-12-13-13-29- 17-9-10-11-11-25-15-19-29-15-15-17-17 |
Субклад R1b1b2/M269
Еще один относительно изученный субклад - это R1b1b2 (ранее R1b1c, M269) (http:// www.familytreedna.com/public/atlantic- r1b1c&fixed_columns=on). На самом деле к этому субкладу относится целый ряд производных субкладов, например, R1b1b2a2c (M153), R1b1b2a2d (M167), R1b1b2a2e (M222), R1b1b2a2g (U152), и остальная часть группы R1b1b2a2, но, как я отмечал выше, каждый субклад состоит из других субкладов, от древности до настоящего времени. И у каждого есть свой общий предок.
В данной выборке подавляющая часть гаплотипов, 83%, относится к субкладу R1b1b2 (M269), остальные - R1b1b2d, R1b1b2e и R1b1b2h, они же R1b1b2a2d, R1b1b2a2e и R1b1b2a2g.
Дерево 67-маркерных гаплотипов этого набора гаплотипов приведено на рис. 6. Видно, что у этой выборки гаплотипов - один общий предок.
Расчет времени жизни общего предка проводился по 25-маркерным гаплотипам (рис. 7).
На все 197 гаплотипов приходится 545 и 1340 мутаций, для 12- и 25- маркерных гаплотипов, соответственно. Это дает 147 и 178 поколений, или 3675 и 4450 лет, соответственно.


Базовый (предковый) гаплотип субклада R1b1b2 (R1b1c) (рис. 7) следующий: 13-24-14-11-11-14-12-12-12-13-13-29- 17-9-10-11-11-25-15-19-29-15-15-17-17 Он в точности равен предковому гаплотипу субклада R1b1b1ag2/S28/U152/R1b1c10 (рис. 5), да и времена жизни общих предков у них почти одинаковы - 4450 и 4375 лет назад, соответственно. От предкового гаплотипа «материнской» гаплогруппы R1b (рис. 1, ветвь справа) их отличает только одна мутация (выделено жирным шрифтом), а на самом деле доля единицы: 13-24-14-11-11-14-12-12-12-13-13-29- 17-9-10-11-11-25-15-19-30-15-15-17-17 Иначе говоря, все эти предковые гаплотипы являются близкородственными, и общие предки жили в пределах нескольких столетий, или даже нескольких поколений, примерно 4200-4600 лет назад. Ни о каких 30 тысяч лет назад, ни о каких кроманьоньцах не может быть и речи. Поскольку баски, гаплогруппа подавляющего большинства которых R1b, часто приводятся в качестве древних представителей R1b, рассмотрим их гаплотипы. Гаплотипы R1b у басков В сети есть «Проект басков» (Basque DNA Project - http://www.familitreedna.com/public/BasqueDNA), в котором приведены 12-маркерные гаплотипы R1b (44 гаплотипа), 25-маркерные (17 гаплотипов) и меньшее количество 37- и 67-маркерных гаплотипов. Среди 12-маркерных гаплотипов нет базовых, и считать мутации по отношению к расчетному базовому гаплотипу нецелесообразно, не зная, происходит ли вся выборка от одного общего предка, или от нескольких. В любом случае, поскольку все 44 12-маркерных гаплотипа содержат 140 мутаций, предварительная оценка дает 173 поколения, или 4325 лет до общего предка. Но эту величину надо проверять. Поэтому было построено дерево 25-маркерных гаплотипов (рис. 8). Видно, что дерево разделяется на две основные ветви, и та, что внизу - более старая ветвь, она отходит от ствола дерева дальше, чем верхняя, более молодая ветвь. Базовые гаплотипы этих двух основных ветвей в 12-маркерном формате следующие (старая и молодая ветви, соответственно): 13-24-14-11-11-14-12-12-12-13-13-29 13-24-14-11-11-14-12-12-12-14-13-30 В 25-маркерном формате более старая баскская ветвь имеет следующий предковый гаплотип: 13-24-14-11-11-14-12-12-12-13-13-29-18-9-10-11-11-25-15-19-28-15-15-17-17 Этот гаплотип на две мутации отличается от предковых гаплотипов субкладов R1b1b2/R1b1c/M269 (рис. 7) и R1b1b2a2g/S28/U152/R1b1c10 (рис. 5) [отмечено жирным шрифтом] 13-24-14-11-11-14-12-12-12-13-13-29-17-9-10-11-11-25-15-19-29-15-15-17-17 и на три и четыре мутации, соответственно, от предковых гаплотипов «материнской» и «дочерней» ветви европейских гаплотипов на рис. 1. 13-24-14-11-11-14-12-12-12-13-13-29- 17-9-10-11-11-25-15-19-30-15-15-17-17 13-24-14-11-11-14-12-12-12-13-13-29-17 -9- 9-11-11-25-15-19-29-15-15-16-17  |
Восемь гаплотипов более старой (нижней) ветви содержат 26 мутаций в первых 12 маркерах, что дает 177 поколений до общего предка. Все 25 маркеров этой ветви содержат 50 мутаций, что дает 161 поколений. Совпадение относительно неплохое. Таким образом, общий предок басков гаплогруппы R1b жил 4425 или 4025 лет назад, по расчетам по 12- и 25-маркерным гаплотипам.
Вторая, более молодая ветвь, содержит только пять гаплотипов, и ее базовый гаплотип следующий (отмечены мутации по сравнению со старой ветвью):
13- 24-14-11-11-14-12-12-12-14-13-30-17-9-10-11-11-25-14-18-29-15-15-16-17
Первые 12 маркеров включают только 7 мутаций, что соответствует примерно 1725 годам до общего предка. Полная последовательность из 25 маркеров имеет 17 мутаций, что соответствует 1675 лет до общего предка. Поэтому можно заключить, что общий предок для более молодой ветви жил примерно 1700±50 лет назад, в 4-м веке нашей эры.
Заметим, что эти два 25-маркерных гаплотипа различаются на 7 мутаций (фактически, без округления, на 5.65 мутаций), что соответствует 143 поколениям, или 3575 лет между двумя общими предками. Это означает, что они не могли произойти друг от друга. Они - две различные линии. ИХ общий предок жил примерно 4600 лет назад. Подобную же цифру мы нашли раньше, анализируя европейские гаплотипы гаплогруппы R1b, и заключив, что общий предок их жил на европейском континенте 4,600±600 лет назад. Эту же цифру дает и анализ общего предка гаплогруппы R1b в Европе (рис. 1).
Таким образом, баски гаплогруппы R1b происходят от того же европейского предка, что и практически все европейские генеалогические линии R1b.
Ген лактозной толерантности у басков
В связи с датировкой возникновения гаплогруппы R1b среди басков, представляет интерес обнаружения у басков гена лактозной толерантности (или гена лактазной персистентности). Эти оба названия означают по сути одно и то же, и относятся к гену Т-13910. Лактоза - это дисахарид, основной углевод в молоке, который метаболизируется на первой стадии под действием фермента лактазы. В биохимии окончание «-оза» соответствует сахарам, углеводам (что суть синонимы), а «-аза» - ферментам, биологическим катализаторам. Лактаза гидролизует, расщепляет лактозу на глюкозу и галактозу, которые легко усваиваются организмом. Если гена Т-13910 в организме нет, или он подавлен, фермент лактаза в организме не образуется, углевод лактоза не усваивается, а вместо этого приводит к резкому дискомфорту у тех, кто пьет молоко. Лактоза тогда разрушается не ферментом, а микрофлорой кишечника с образованием большого количества газов (поскольку лактозы в молоке много), что приводит к тому, что «живот пучит». Природа сделала так, что лактаза образуется только при рождении младенцев, и этот фермент активен только додостижения младенцем года-двух, после чего ген дезактивируется, перестает работать.
Но в ходе развитии молочного животноводства у древних народов, у кого-то в генах произошла такая случайная мутация, которая привела к появлению активного гена Т-13910, причем активного всю жизнь его носителя. Естественно, это дало преимущества мутированной особи в отношении естественного отбора, его носители пили молоко без вреда для организма, и мутация закрепилась в последующих поколениях. Исследования этого гена привели к региону его первоначального появления - либо в западном Предуралье, 6600-4800 лет назад (Anthony, 2007) , либо на Кавказе или к северу от Кавказа, между 5 и 12 тысяч лет назад, где наблюдается наиболее высокое разнообразие мутаций соответствующего сегмента ДНК. Значительное разнообразие мутаций этого гена было выявлено также в Азии, на восточной стороне Уральских гор, а также в Средней Азии.
Недавние исследования показали, что этот ген лактозной устойчивости находится у 92% басков (Enattah et al, 2007). Более того, этот же ген распространен в Европе и западной Азии, он обусловлен единичной мутацией, и эта мутация является уникальной и окружена в соответствующем фрагменте ДНК характерной «подписью» нуклеотидов. Так что ген один и тот же и в Европе, и в Азии.
Эти данные согласуются с теми, что современные баски являются потомками мигрантов в составе рода R1b в Испанию и Францию 43004600 лет назад, или в любом случае не раньше 5-12 тысяч лет назад, из Азии или с Кавказа. Более того, изучение ДНК останков древних европейцев показало, что ген Т-13910 не наблюдался в них ранее, чем 7000-7800 лет назад. Это все показывает, что современные баски отнюдь не происходят от народов, населявших Европу 30 тысяч лет назад. Во всяком случае, не от кроманьонцев.
Поскольку этот ген расположен не в Y-хромосоме, то он передавался через женщин носителям других мужских гаплогрупп, и таким образом распространился по разным гаплогруппам. В отношении исходного региона возникновения гена Т-13910 исследователи склоняются к двум регионам - Средней Азии, и территории к северу от Кавказа и западу от Урала, то есть южным степям России. Оба региона могут иметь отношение к возникновению гаплогруппы (мутации) R1b или быть на путях миграции рода R1b1 на Кавказ и в Европу.
Гаплотипы R1b1b2 у жителей Буковины, северные Карпаты
Буковина - старое название местности на северо-востоке Карпат, на стыке Украины и Румынии, со стороны Украины - Черновицкая область. Город Черновцы и есть исторический центр Буковины. В рамках археологии - часть территории трипольской культуры. В недавней работе (Клёсов, 2008а) показано, что носители гаплогруппы R1a1 появились там около 6000 лет назад, причем те носители, потомки которых живут в наше время. Точнее, по 25-маркерным гаплотипам время жизни общего предка R1a1 было 5975 лет назад, по менее точным 12-маркерным гаплотипам - 4675 лет назад.
Посмотрим на гаплотипы Буковины гаплогруппы R1b1b2. Дерево 12- маркерных гаплотипов показано на рис. 9.
Во всех 19 гаплотипах в 12-маркерном формате содержится 72 мутации, что дает 213 поколений, то есть 5325 лет до общего предка. В девяти 25- маркерных гаплотипах содержится 64 мутации, что дает до общего предка 187 поколений, или 4675 лет. Таким образом, по сравнению с гаплотипами R1a1 из того же региона 25-маркерные гаплотипы R1b «моложе» на 1300 лет, и 12-маркерные гаплотипы «старше» на 650 лет. В целом R1b гаплотипы Буковины старше центрально- и западноевропейских, что согласуется с их миграцией в Европу с востока.

Гаплотипы R1b1b2-M269 на Сардинии 8-маркерные гаплотипы жителей этого средиземноморского острова недавно были опубликованы (Contu et al, 2008). В статье отмечалось, что остров был населен уже в неолитические времена, и во времена бронзового века численность населения стала возрастать, достигнув 300 тысяч человек 3600-2200 лет назад. Естественно, состав того населения по гаплогруппам неизвестен, но может быть хотя бы примерно реконструирован по гаплогруппам современников. Из 376тестированных человек 64 (17%) оказались принадлежащими к гаплогруппе R1b1b2 (снипы М89, М9, М173, М269). В другой выборке тестировали 930 человек, и 158 (те же 17%) из них оказались с гаплогруппой R1b1b2. Авторы применили некритические методы расчетов (с использованием «коэффициента Животовского», упомянутого выше) и получили, что общий предок носителей этой гаплогруппы на Сардинии жил 22800 лет назад. Это, конечно, фантазийная цифра, но какая ближе к действительности? Для ответа на этот вопрос были построены деревья 8-маркерных гаплотипов (рис. 10 и 11). Дерево на рис. 10 более древнее, в него входят гаплотипы «родительских» гаплогрупп K-R и R1, а также гаплогруппы R1b1a. 10 гаплотипов группы K-R имеют 50 мутаций от расчетного предкового гаплотипа 13-23-15-10-14-16-X-Y-Z-13-W-30 что соответствует 424 поколениям, или 10600 лет до общего предка. Четыре гаплотипа гаплогруппы R1 принадлежали, видимо, довольно близким родственникам, так как они практически не отличались друг от друга 13-23-15-11-14-14-X-Y-Z-14-W-31 На все 32 маркера была всего лишь одна мутация, что помещает их общего предка в пределах 15 поколений, или около 400 лет от настоящего времени. Но разница в предковых гаплотипах K-R и R1, приведенных выше, помещает ИХ общего предка на 10800 лет назад, то есть почти совпадает с общим предком для сводной гаплогруппы K-R на Сардинии.  |
13-24-15-10-11-12-X-Y-Z-13-W-28
имел всего 10 мутаций на восьми гаплотипах, что дает 2000 лет до общего предка. Как будет показано ниже, этот предковый гаплотип отличается от предкового гаплотипа гаплогруппы R1b1c на Сардинии на 8 мутаций на 8 маркерах, что помещает ИХ общего предка на 15 тысяч лет назад. Кстати, наличие нескольких гаплотипов гаплогруппы R1a1 на Сардинии позволяет оценить время жизни их общего предка, что составляет 4400 лет, как и по всей Европе, за исключением Балкан (Клёсов, 2008а).
Для рассмотрения дерева гаплотипов R1b1b2 гаплотипы K-R, R1 и R1b1a были удалены, и оставшиеся гаплотипы (снип М269) приведены на рис. 11.
Видно, что полученное дерево состоит из нескольких ветвей гаплотипов: серия из семи идентичных гаплотипов на «стволе» дерева, отдельная ветвь из 12 гаплотипов справа вверху, ветвь из 16 гаплотипов слева вверху, и остальная часть дерева.
Разберем эти ветви по частям.
Семь одинаковых гаплотипов, образующих короткую «гребенку» на вершине дерева, имеют следующий состав:
13-24-14-11-11-14-X-Y-Z-13-W-29
Это - классический «Атлантический модальный гаплотип», который в полной 12-маркерной записи выглядит так:
13-24-14-11-11-14-12-12-12-13-13-29
Ясно, что эти гаплотипы - от недавнего предка, удаленного по времени от остальных гаплотипов на дереве гаплотипов на рис. 11.
Для рассмотрения дерева гаплотипов R1b1b2 гаплотипы K-R, R1 и R1b1a были удалены, и оставшиеся гаплотипы (снип М269) приведены на рис. 11.
Видно, что полученное дерево состоит из нескольких ветвей гаплотипов: серия из семи идентичных гаплотипов на «стволе» дерева, отдельная ветвь из 12 гаплотипов справа вверху, ветвь из 16 гаплотипов слева вверху, и остальная часть дерева.
Разберем эти ветви по частям.
Семь одинаковых гаплотипов, образующих короткую «гребенку» на вершине дерева, имеют следующий состав:
13-24-14-11-11-14-X-Y-Z-13-W-29
Это - классический «Атлантический модальный гаплотип», который в полной 12-маркерной записи выглядит так:
13-24-14-11-11-14-12-12-12-13-13-29
Ясно, что эти гаплотипы - от недавнего предка, удаленного по времени от остальных гаплотипов на дереве гаплотипов на рис. 11.

12 гаплотипов правой верхней ветви имеют 25 мутаций от базового гаплотипа 13-24-14-10-11-14-X-Y-Z-13-W-29 который отличается всего на одну мутацию от АМГ на том же дереве. Эти 25 мутаций соответствуют 141 поколению, или 3525 лет до общего предка. 16 гаплотипов левой верхней ветви имеют 28 мутаций от базового гаплотипа 13-24-14-10-11-15-X-Y-Z-13-W-29 который отличается на две мутации от АМГ на том же дереве, и всего на одну мутацию от предыдущего базового гаплотипа. Эти 28 мутаций соответствуют 116 поколениям или 2900 лет до общего предка. Наконец, если объединить все ветви (что, строго говоря, не является правильным, поскольку ветви разного размера), то окажется, что все 64 гаплотипа содержат 180 мутаций, приводя к 201 поколению, или 5025 лет до общего предка всех носителей этих гаплотипов. Эта величина имеет условную надежность, но показывает, что она совершенно далека от величины 22800 лет, «определенную» авторами статьи (Contu et al, 2008) с использованием скорости мутации 0.00069 на маркер, то есть в 3.1 раза ниже, чем принятые в научном сообществе, причем с отбрасыванием двух маркеров из восьми. Гаплотипы R1b в Сицилии В работе (Di Gaetano et al, 2008) приведены всего девять 10-маркерных гаплотипов группы R1b с Сицилии. Этого количества недостаточно, чтобы строить дерево гаплотипов, но достаточно для проведения предварительной оценки времени жизни общего предка гаплогруппы на острове. Эти гаплотипы имеют следующий базовый гаплотип 13-24-14-11-11-14-X-Y-12-13-13-29 который идентичен гаплотипу относительно недавнего предка на Сардинии 13-24-14-11-11-14-X-Y-Z-13-W-29 и идентичен Атлантическому модальному гаплотипу 13-24-14-11-11-14-12-12-12-13-13-29 в пределах определенных аллелей. Все девять гаплотипов имеют 25 мутаций, что соответствует 143 поколениям, или 3600 лет до общего предка гаплогруппы R1b на Сицилии. Это - обычные европейские величины для гаплогруппы R1b. Гаплотипы R1b1b2 в Ливане Недавно были опубликованы 44 гаплотипа гаплогруппы R1b1b2 (бывшая R1b1c) из Ливана, наряду с десятью R* гаплотипами и тремя гаплотипами R1b1a из того же региона (Zalloua et al, 2008). Общее дерево этих гаплотипов дано на рис. 12. Видно, что только три гаплотипа группы R* (под номерами 09, 10, 11) и все три гаплотипа R1b1a (14, 15, 16) выбиваются из общей структуры дерева. Остальные семь гаплотипов R* встроены в общую структуру, и даже являются частью относительно «молодых» ветвей. При последующих расчетах я использовал только девять маркеров из 11, приведенных авторами, так как маркеры DYS437 и DYS438 принадлежат другим панелям, и могут внести неопределенность в расчеты. Все десять гаплотипов R* сходятся к их предковому (базовому) гаплотипу, который идентичен, как будет показано ниже, базовому гаплотипу R1b1b2 12- 24-14-10-X-Y-Z-12-12-13-13-29 Этот гаплотип отклоняется на две мутации от Атлантического модального гаплотипа: 24-14-11-11-14-12-12-12-13-13-29  |
Все 10 гаплотипов группы R* содержат 37 мутаций от базового, что соответствует 262 поколениям, или 6550 годам до общего предка этих гаплотипов.
Три гаплотипа группы R1b1a, конечно, недостаточны для расчетов, поэтому была проведены просто приблизительная оценка их «возраста» относительно общего предка с гаплотипом (ориентировочно)
13-24-15-10-X-Y-Z-13-12-13-13-29
На три гаплотипа приходится 8 мутаций, что примерно соответствует 4400 лет до общего предка.
Перейдем к гаплотипам группы R1b1b2 (рис. 13).

| Видно, что выборка содержит несколько линий недавних предков (плоские ветви на рис. 13), но в целом не содержит сепаратных ветвей. Это означает, что для дерева можно принять одного общего предка. Его гаплотип - тот самый предковый гаплотип R*, приведенный выше 12-24-14-10-X-Y-Z-12-12-13-13-29 и который отклоняется на две мутации от АМГ. Все 44 гаплотипа содержат 137 мутаций, что дает 212 поколений, или 5300 лет до общего предка гаплогруппы R1b1b2 в Ливане. Это - несколько выше, хотя и не намного, возраста общих предков этой гаплогруппы в Европе. Чтобы определить, действительно ли гаплогруппа R1b1b2 в Ливане старше европейских, применим два критерия - сравним с возрастом гаплогруппы R1a1 в том же регионе, и сравним с возрастом европейских гаплотипов R1b (M343), представленных в той же статье (Zalloua et al, 2008), по их 12- и 9-маркерным гаплотипам.  |
15 гаплотипов R1a1 из Ливана расходятся по двум совершенно разным ветвям (рис. 14). Справа - относительно молодая ветвь, в ней всего четыре гаплотипа, содержащих пять мутаций от ориентировочного предкового гаплотипа R1a1 в Ливане: 13-24-15-11-X-Y-Z-12-10-14-11-31 Это дает «возраст» общего предка данных четырех гаплотипов всего лишь 75 поколений назад, то есть 1875 лет. Наша эра, 2-й век. Более старая, левая ветвь на рис. 14, имеет базовый гаплотип 13-25-16-10-X-Y-Z-12-10-13-11-30 и содержит 29 мутаций на 9 гаплотипов. Это соответствует 221 поколению, или 5525 лет до общего предка. Сам гаплотип идентичен на данных маркерах типичному европейскому предковому гаплотипу R1a1 13-25-16-10-11-14-12-12-10-13-11-30 который в свою очередь в точности соответствует восточнославянскому базовому (предковому) гаплотипу. В любом случае, выборки ливанских гаплотипов R1b1b2 и R1a1 имеют общих предков практически одинакового возраста, 5300 и 5525 лет, соответственно. Проверим это на других системах. На рис. 15 приведено дерево 12-маркерных европейских гаплотипов, построенное по данным, приведенным в той же работе (Zalloua et al, 2800) . Видно, что дерево практически симметричное, следовательно, происходит от одного общего предка. Все 112 гаплотипов содержат 334 мутации, что соответствует 160 поколениям, или 4000 лет до общего предка. Те же 112 гаплотипов, но 9-маркерные, содержат 258 мутаций, что соответствует 148 поколениями, или 3700 лет до общего предка.  |
Иначе говоря, расчет по 9-маркерным гаплотипам в данном случае даже несколько занижает расстояние до общего предка, и указанные 5300 (R1b1b2) и 5525 (R1a1) лет в Ливане могут быть даже выше на несколько сот лет, например, 5700 и 6000 лет до общего предка. Гаплотипы R1b1b2 в Анатолии 84 десятимаркерных гаплотипа R1b1b2 (M269) были приведены в статье (Cinnioglu et al, 2004). Среди них был маркер DYS#A7.2 (DYS461), который не числится в первых 37 маркерах в системе записи FTDNA, и поэтому был мной исключен из рассмотрения. Дерево 9-маркерных гаплотипов приведено на рис. 16.  |
Из 84 гаплотипов восемь относятся к отдельным гаплогруппам или субкладам (гаплотипы 001 и 002 принадлежат гаплогруппе R1b1/P25, 079-082 представляют гаплогруппу R1b1b/M73, 083-084 относятся к исходной гаплогруппе R1b/M343), и не были включены в расчеты. Видно, что дерево на рис. 16 состоит из нескольких выраженных ветвей. На вершине дерева находятся семь идентичных гаплотипов 12-24-14-11-X-Y-Z-12-12-13-13-29 определенно от относительно недавнего общего предка. Этот гаплотип отклоняется всего лишь на одну мутацию от предкового ливанского гаплотипа той же гаплогруппы R1b1b2 12- 24-14-10-X-Y-Z-12-12-13-13-29, на одну и две мутации от предковых гаплотипов R1b1b2 Сардинии 13-24-14-11-11-14-X-Y-Z-13-W-29 13-24-14-10-11-15-X-Y-Z-13-W-29 на одну мутацию от предкового гаплотипа Сицилии (идентичного АМГ) 13-24-14-11-11-14-X-Y-12-13-13-29 и на одну мутацию от усеченного Атлантического модального гаплотипа (АМГ): 13-24-14-11-11-14-12-12-12-13-13-29 Ветвь справа вверху на рис. 16 состоит из 13 гаплотипов R1b1b2, с предковым гаплотипом 13-24-14-11-X-Y-Z-12-12-13-13-29 Это - опять усеченный Атлантический модальный гаплотип. Все 13 гаплотипов содержат 27 мутаций, что дает 132 поколения, или 3300 лет до общего предка. Широкая ветвь из 33 гаплотипов R1b1b2 в нижней части дерева на рис. 15 содержит 99 мутаций, что дает 200 поколений, или 5000 лет до общего предка с базовым гаплотипом 12-24-14-10-X-Y-Z-12-12-13-13-29 Наконец, все 76 R1b1b2 гаплотипов содержат 207 мутаций, что дает 180 поколений, или 4500 лет до общего предка. Эта величина занижена, поскольку ее «тянут вниз» гаплотипы от недавних предков с малым количеством мутаций. Именно поэтому необходимо строить деревья и разделять гаплотипы на ветви. Подводя итог данному разделу, гаплотипы R1b1b2 Анатолии имеют древнейшего общего предка, который жил 5000 лет назад, примерно тогда же, когда и в Ливане (5300 лет назад). Расчеты времени жизни общего предка гаплогруппы R1a1 на тех же территориях дали 4500 и 5525 лет, соответственно. Иначе говоря, представители родов R1a1 и R1b1b2 жили на этих территориях в одно и то же время. Гаплотипы R1b в Армении Выборка из 238 гаплотипов армян гаплогруппы R1b была представлена в работе (Weale et al, 2001). Она включала гаплотипы из шести регионов Армении, Карабаха, Ирана, и других районов армянской диаспоры в мире. Рассмотрение нами этих гаплотипов показало, что часть из них относятся к древним, а часть - к относительно «молодым» предкам. Иначе говоря, все эти выборки представляют неоднородную смесь популяций, и для всех расчеты по остаточным гаплотипам и по мутациям дают весьма различные результаты. Тем не менее, во всех шести регионах базовый гаплотип был один и тот же: 12-24-14-11-X-X-X-12-X-X-13-X Он отличается на одну мутацию от «Атлантического модального гаплотипа» 12-24-14-10-X-Y-Z-12-12-13-13-29, Если не проводить разделение на отдельные ветви гаплотипов, и считать общих предков по регионам, то, например, для Иранского региона число поколений до «общего предка», рассчитанное по остаточным базовым гаплотипам (Клёсов, 2008b) и по мутациям оказалось равно ln(18/4)/0.00877 = 214 и 244, соответственно. Для Карабаха это равно 185 и 206 поколений. Для Восточной Турции - 237 и 286 поколений. Для Северной Армении - 249 и 291 поколение. Для Южной Армении - 151 и 178 поколений. Самое высокое число поколений оказалось для Араратского региона - 258 и 400 поколений, то есть, формально (и неправильно) говоря, 6,450 и 10,000 лет до общего предка. Дело в том, что при смеси общих предков в выборке первая цифра обычно оказывается завышенной, а вторая - заниженной. При разделении предков эти цифры обычно расходятся шире. Насколько шире расходятся - зависит от ряда факторов, в особенности от того, сколько потомков в выборке от каждого общего предка, и сколько до него поколений от настоящего времени. Для того, чтобы разобраться с этой проблемой, было построено дерево для 52-х гаплотипов для объединенной выборки Араратского региона и Северной Армении (рис. 17) Дерево подразделилось на пять ветвей. Две самые древние, но очень малочисленные ветви расположены слева внизу и справа вверху на рис. 17. Шесть гаплотипов первой содержат 18 мутаций, что формально соответствует 491 поколению, или 12300 лет до общего предка. Другая ветвь дала 13 мутаций на четыре гаплотипа, что соответствует 553 поколениям, или 13800 лет до общего предка. Третья, широкая ветвь в нижней правой части на рис. 17, состоит из 21 гаплотипа и содержит 36 мутаций, что соответствует 235 поколениям, или примерно 5900 лет до общего предка. Четвертая, плоская ветвь в левом верхнем секторе, из 10 гаплотипов, содержит 16 мутаций, что соответствует 216 поколениям, или 5400 лет до общего предка. Наконец, пятая ветвь оказалась совсем недавней, в ней были только 11 идентичных гаплотипов, поэтому возраст посчитать было нельзя. Примерно несколько сот лет до общего предка.  Для сравнения, общий предок армянских R1a1 гаплотипов жил 4400 лет назад (Клёсов, 2008а). Ясно, что большинство армянских общих предков гаплогруппы R1b значительно старше западно-европейских, которые жили в основном между 3400 и 4400 годами назад. Гаплотипы R1b на Балканах В частном порядке я получил от д-ра Marijana Pericic серию из 36 гаплотипов гаплогруппы R1b, собранных на разных территориях Балкан, побережья Адриатического моря, и островах Адриатики. Дерево гаплотипов показано на рис. 18.  Видно, что дерево гаплотипов содержит несколько ветвей, что соответствует нескольким общим предкам. Можно выделить четыре явно разные ветви, соответствующие четырем генеалогическим линиям. Одна из них совсем недавняя, из 15 гаплотипов, включает 12 базовых гаплотипов, и содержит всего три мутации. Это дает по остаточным базовым гаплотипам ln(15/12)/0.0173 = 13 поколений, и по мутациям 3/15/0.0173 = 12 поколений. Таким образом, молодая ветвь, в количестве одной трети от всех R1b, имеет общего предка, который жил всего 300-325 лет назад, в конце 17-го - начале 18-го веков. Почти все они - жители острова Корсула в Адриатике. Базовый гаплотип у них следующий: 13-24-14-11-11-11-X-Y-Z-13-13-29 Он на три мутации отходит от известного Атлантического модального гаплотипа (АМГ): 13-24-14-11-11-14-12-12-12-13-13-29 Вторая ветвь, справа внизу на рис. 18, содержит 11 или 14 гаплотипов, в зависимости от того, как ее рассматривать. Базовый гаплотип при этом не меняется: 12- 24-14-11-11-15-X-Y-Z-13-13-29 Здесь - две мутации в сторону от АМГ В этой ветви 26 мутаций при 11 гаплотипах, или 31 мутация при 14 гаплотипах. Это дает соответственно 157 или 146 поколений, то есть 3,900 или 3,700 лет до общего предка. Можно записать 3,800±100 лет. Третья ветвь, слева внизу, из 6 гаплотипов, содержит 17 мутаций от базового гаплотипа 13- 24-14-11-11-11-X-Y-Z-14-13-29 и отстоит на четыре мутации от АМГ. Возраст до общего предка этой ветви составляет 4,900 лет. Наконец, самая древняя ветвь, как и видно из дерева гаплотипов, состоит из 7 гаплотипов и содержит 21 мутацию. Это дает 208 поколений, или 5200 лет до общего предка, что немногим отличается от других цифр для возраста общего предка R1b в Европе. Более того, этот возраст в данном случае держится только на одном гаплотипе, номер 042 на рис. 18. Если его убрать, то останется 15 мутаций на 6 гаплотипах, и это даст вполне умеренные для Европы 168 поколений, то есть примерно 4,200 лет до общего предка. Но такие разбросы, к сожалению, это общие недостатки малых выборок. Шведские гаплотипы гаплогруппы R1b1b2 Набор из 383 девятимаркерных гаплотипов Швеции, включавший 76 гаплотипов R1b1b2, был опубликован в работе (Karlsson et al, 2006). Базовый гаплотип R1b1b2 в Швеции следующий: 13-24-14-11-11-14-X-Y-Z-13-13-29 Это - классический Атлантический модальный гаплотип. Все 76 шведских гаплотипов гаплогруппы R1b1b2 включают 7 базовых гаплотипов и содержат 187 мутаций. Без поправки на возвратные мутации это дает ln(76/7)/0.0173 = 138 поколений, и 187/76/0.0173 = 142 поколений до общего предка. Опять мы видим почти одинаковые величины, что указывает на одного общего предка в выборке. Поскольку число поколений превышает сотню, необходимо ввести поправку на возвратные мутации, которые в данном случае увеличат возраст предка примерно на 16%, а именно до 165 поколений, то есть до 4,125 лет до общего предка. Это - совершенно обычная цифра для европейских гаплотипов гаплогруппы R1b1b2. 
|

Сравним два дерева гаплотипов R1b, общего по Европе (рис. 1) и фламандское (рис. 20). Разница принципиальная. У фламандского - явно один общий предок. По доле остаточных базовых гаплотипов число поколений до общего предка равно ln(64/2)/0.0218 = 159±20. Поскольку все 64 гаплотипа содержат 199 мутаций, то 199/64/0.0218 = 143 поколения. Погрешность в логарифмическом варианте рассчитывалась исходя из того, что если бы в выборке по статистической случайности оказалось не два, а три базовых гаплотипа, то ln(64/3)/0.0218 = 140 поколений. Согласование числа поколений до общего предка, рассчитанного по числу базовых гаплотипов и по числу мутаций показывает, что общий предок был один для всех 64 человек, и наши методы расчета полностью применимы.
С поправкой на возвратные мутации получим, что общий предок фламандцев гаплогруппы R1b из данной выборки жил 166 поколений, или 4,150 лет назад. Это близко к времени жизни общего предка гаплогруппы R1b1b2 в Швеции (4,125 лет назад), общим предком R1b в Европе, рассчитанным по 12-маркерным гаплотипам (3,375 лет назад), и с прочими данными для R1b на разных европейских территориях.
Гаплотипы R1b1b2 и R1b1b1/M73 из Средней Азии
Ниже приведены четырнадцать 25-маркерных гаплотипов среднеазиатского происхождения. Это гаплотипы узбеков, таджиков, тувинцев, уйгур, казахов, полученные непосредственно от работающих там исследователей.

| (список гаплотипов приведен в ссылке http://dnatree.m/index.php?name=Forams&file=viewtopic&p=18889#188 24) Видно, что этот список гаплотипов неоднородный, и представляет две разные серии, каждая со своим общим предком. Этому соответствует нумерация гаплотипов от 001 до 006, и от 101 до 108. На дереве гаплотипов (рис. 21) они расходятся по совершенно разным ветвям.  |
Все 6 гаплотипов в сумме имеют 16 мутаций на первых 72 маркерах (то есть на первой 12-маркерной панели), у них мутированы 22% маркеров. Это - хорошая статистика, предок довольно древний. Во всех 25- маркерных панелях у них 25 мутаций. По числу мутаций получаем - для 12-маркерных гаплотипов 141 поколение, или 3525 лет до общего предка, и для 25-маркерных 102 поколения, или 2550 лет до общего предка. В среднем 122±28 поколений, или 3100±700 лет до общего предка. Это - довольно обычные времена жизни общего предка R1b в Европе, и можно было бы полагать, что у приведенных шести гаплотипов он имеет европейское происхождение. Однако это не так. По сравнению с «модальным атлантическим гаплотипом» (АМГ) 13 24 14 11 11 14 12 12 12 13 13 29 17 9 9(10) 11 11 25 15 19 29(30) 15 15 16(17) 17 определенный выше базовый гаплотип имеет 8 мутаций на первых 12 маркерах, и 15-18 мутаций на всех 25 маркерах (соответствующие аллели выделены жирными цифрами). Это - очень большое расстояние во времени, и оно соответствует примерно 580 поколений на первых 12 маркерах, и 500-650 поколений на 25-маркерных гаплотипах. А поскольку мы знаем, что западноевропейский АМГ имеет общего предка 185 поколений, или 4625 лет назад (см. выше), то можно рассчитать, что общий предок западноевропейского АМГ и среднеазиатского предкового гаплотипа, приведенного выше, жил [122+185+(500-650)]/2 = от 400 до 480 поколений назад, или между 10000 и 12000 лет назад. Если усреднить, получится 11000±1400 лет назад. Он и был общим предком для европейских и приведенных выше шести азиатских представителей гаплогруппы R1b. У левой ветви на рисунке базовый, или предковый гаплотип следующий: 13 19 14 11 13 13 12 12 14(13) 14 13 30 17 9 9 11 11 23 15 19 33 12 15 15 16 Общий предок всех восьми человек с этими гаплотипами жил совсем недавно по историческим меркам. Все восемь гаплотипов в сумме имеют всего пять мутаций на первых 96 маркерах (то есть на первой 12- маркерной панели), у них мутированы всего 5% маркеров. Во всех 25- маркерных панелях у них всего 18 мутаций. Поскольку из восьми 12-маркерных гаплотипов четыре идентичны, то есть сохраняют предковый гаплотип, то расчеты показывают: ln(8/4)/0.022 = 31.5 поколений до общего предка. По числу мутаций в тех же восьми гаплотипах получаем 5/8/0.022 = 28.4 поколения до общего предка. Эти величины весьма близки, и в среднем дают 30±2 поколений. Такое совпадение показывает, что предок у данных восьми человек был действительно один, общий для всей серии. Во всех 25-маркерных гаплотипах содержится 18 мутаций, поэтому 18/8/0.046 = 49 поколений, или 1225 лет до общего предка. Подобные разнобои между двумя панелями встречаются обычно тогда, когда количество мутаций в панели меньше 8-10% от числа маркеров. Поэтому примем, что общий предок для всех 8 человек жил 1225 лет назад, то есть примерно в 8-м веке нашей эры. Самое интересное здесь то, что предковый гаплотип имеет аллель 19 во втором слева маркере (DYS390, по принятой номенклатуре), в то время как «классический» западноевропейский «атлантический модальный гаплотип» (АМГ) имеет там аллель 24. Пять мутаций - это не шутка, это многие тысячи лет разницы в эволюции гаплотипа. Вот так, с отмеченными мутациями, выглядит западноевропейский АМГ: 13 24 14 11 11 14 12 12 12 13 13 29 17 9 9(10) 11 11 25 15 19 29(30) 15 15 16(17) 17 Отличие на 12-маркерной панели составляет 11 или 12 мутаций, на всей 25-маркерной панели - от 19 до 25 мутаций. Это - огромное расстояние во времени, и соответствует примерно от 960 до 1100 поколений. А поскольку мы знаем, что западноевропейский АМГ имеет общего предка примерно 185 поколений, или 4625 лет назад (см. выше), то можно рассчитать, что общий предок западноевропейского АМГ и среднеазиатского предкового гаплотипа, приведенного выше, жил [49+185+(960-1100)]/2 = от 600 до 680 поколений назад. Таким образом, разумно будет принять, что общий предок R1b жил примерно 16000±1400 лет назад. Он и был предком для европейских и азиатских представителей гаплогруппы R1b. На самом деле азиатский гаплотип с DYS390 = 19 принадлежит гаплогруппе R1b1b1 (M73). Он довольно широко распространен от Турции до Сибири. Как показано выше, он весьма далеко отстоит по мутациям (и, соответственно, по времени) от обычного европейского гаплотипа R1b1b2 (M269). Посмотрим, наконец, а как далеко разнесены во времени два приведенных выше азиатских предковых гаплотипа R1b: 13 22 14 11 13 17 12 12 12 13 13 30 15 9 9 11 11 23 15 20 29 12 15 15 17 12 19 14 11 13 13 12 12 14(13) 14 13 30 17 9 9 11 11 23 15 19 33 12 15 15 16 Между ними - 9-10 мутаций на 12-маркерной панели, и 17-18 мутаций на 25-маркерных предковых гаплотипах. Это - 690-810 поколений на 12- маркерной панели, и 590-650 поколений на 25-маркерной панели, или 690±90 поколений. Поскольку мы знаем, что обладатели этих базовых гаплотипов жили примерно 122 и 49 поколений назад, то ИХ общий предок жил (122+49+690)/2 = 430 поколений, или 10800 лет назад. Это соответствует значению 11000±1400 лет назад, приведенному выше. Заключение Итак, общий предок гаплогруппы R1b в Азии жил примерно 16 тысяч лет назад. Потомки его ушли долгим путем в Европу, перед этим 1200014000 лет назад жили в Армении, на что указывают армянские гаплотипы R1b (см. выше), и примерно 4000-4500 лет назад пришли в Европу. Тем временем оставшаяся азиатская ветвь примерно 11 тысяч лет назад в свою очередь расщепилась на две ветви, обе среднеазиатские. Они сейчас обнаружены по их относительно недавним предкам, жившим примерно 3100 и 1220 лет назад. Гаплотипы этих предков описаны выше. Возможно, обе популяции прошли бутылочное горлышко три тысячелетия и тысячелетие назад, и то, что мы видим - это две выживших генеалогических линии, довольно далеко разошедшиеся за 10-12 тысяч лет. Но они продолжают вести линию их древнего общего предка, основавшего род R1b. Можно предположить, что с Кавказа носители гаплогруппы R1b отправились сначала на север, вслед за уходящими ледниками. Много позже они оставили следы своего пребывания в виде археологических культур - возможно, культуры шнуровой керамики, она же культура боевых топоров, она же культура одиночных могил. У археологов пока нет ответа на вопрос, к каким родам принадлежали эти культуры. Мы не можем исключить, что ямная или/курганная культуры также были оставлены родом R1b. Похоже, пути движения родов R1b и R1a1 пересеклись 6000-4000 лет назад, первые в целом передвигались на запад, вторые - на восток. Территория Восточной Европы хранит память о том времени в виде большого количества древних черепов, раздробленных каменными боевыми топорами. Типичные представители гаплогруппы R1b - кельты, которые появились в Западной Европе 3500-4500 лет назад. Кстати, кельты - имя собирательное и впервые примененное в его современном значении не так давно, в начале 18-го века, Эдвардом Лайдом, директором Ашмолеанского музея в Оксфорде. Путешествуя по делам музея, он обратил внимание на сходство языков уэльсцев, корнишей, бретонцев, ирландцев, шотландских галлов, и древних галльских языков. Он и объединил эти языки под общим, придуманным им именем кельтских языков. Хотя само имя кельты упоминал еще Юлий Цезарь в книге «Записки о галльской войне», как синоним галлов. Таблица ниже представляет полученные в настоящей работе данные по временам жизни общих предков.  |


Литература Клёсов, А.А. Откуда появились славяне и «индоевропейцы» и где их прародина? Ответ дает ДНК-генеалогия. Вестник Российской Академии ДНК-генеалогии. 1, 400-477 (2008a) Клёсов, А.А. Основные положения ДНК-генеалогии (хромосома Y), скорости мутаций, их калибровка и примеры расчетов. Вестник Российской Академии ДНК-генеалогии. 1, № 2, 252-348 (2008b). Anthony, D.W. The Horse, The Wheel, and Language: How Bronze-Age Riders from the Eurasian Steppes Shaped the Modern World. Princeton University Press, 2007, p. 326. Campbell, K.D. Geographic patterns of haplogroup R1b in the British Isles. J. Genetic Genealogy 3, 1-13 (2007) Cinnioglu, C., King, R., Kivisild, T., Kalfoglu, E., Atasoy, S., Cavalleri, G., Lillie, A.S., Roseman, C.C., Lin, A.A., Prince, K., Oefner, P.J., Shen, P., Semino, O., Cavalli-Sforza, L.L. and Underhill, P,A. Excavating Y- chromosome haplotype strata in Anatolia. Hum. Genet. 114, 127-148 (2004) Contu, D., Morelli, L., Santoni, F., Foster, J.W., Francalacci, P. And Cucca, F. Y-Chromosome based evidence for pre-neolithic origin of the genetically homogeneous but diverse Sardinian population: inference for association scans. PLoS ONE, 1, 1-8 (2008). Derenko, M., Malyarchuk, B., Denisova, G., Wozniak, M., Grzybowski, T., Dambueva, I. and Zakharov, I. Y-chromosome haplogroup N dispersals from south Siberia to Europe. J. Hum. Genet. 52, 763-770 (2007) Di Gaetano, C., Cerutti, N., Crobu, F., Robino, C., Inturri, S., Gino, S., Guarrera, S., Underhill, P.A., King, R.J., Romano, V., Cali, F., Gasparini, M., Matullo, G., Salerno, A., Torre, C., and Piazza, A. Differential Greek and northern African migrations to Sicily are supported by genetic evidence from the Y chromosome. Eur. J. Human Genetics, August 2008, preprint. Enattah, N. S., Trudeau, A., Pimenoff, V., Maiuri, L., Auricchio, S., Greco, L., Rossi, M., Lentze, M., Seo, J.K., Rahgozar, S., Khalil, I., Alifrangis, M., Natah, S., Groop, L., Shaat, N., Kozlov, A., Verschubskaya, G., Comas, D., Bulayeva, K., Mehdi, S.Q., Terwilliger, J.D., Sahi, T., Savilahti, E., Perola, M., Sajantila, A., Jarvela, I., and Peltonen L. Evidence of still-ongoing convergence evolution of the lactase persistence T-13910 alleles in humans. Am J Hum Genet. 81, 615-25 (2007). Gayden, T., Cadenas, A.M., Regueiro, M., Singh, N.B., Zhivotovsky, L.A., Underhill, P.A., Cavalli-Sforza, and Herrera, R. J. The Himalayas as a directional barrier to gene flow. Amer. J. Human Genetics, 80 (2007) Karlsson, A.O., Wallerstrom, T., Gotherstrom, A. and Holmlund, G. Y- chromosome diversity in Sweden - A long-time perspective. Europ. J. Human Genetics, 14, 963-970 (2006) Martinez, L., Underhill, P.A., Zhivotovsly, L.A., Gayden, T., Moschonas, N.K., Chow, C.-E. T., Conti, S., Mamolini, E., Cavalli-Sforza, L.L. and Herrera, R.J. Paleolitic Y-haplogroup heritage predominates in a Cretan highland plateau. Eur. J. Human Genetics, 15, 485-493 (2007) Mertens, G. Y-Haplogroup frequencies in the Flemish population. J. Genetic Genealogy 3, 19-25 (2007) Sengupta, S., Zhivotovsky, L.A., King, R., Mehdi, S.Q., Edmonds, C.A., Chow, C.-E. T., Lin, A.A., Mitra, M., Sil, S.K., Ramesh, A., Rani, M.V.U., Thakur, C.M., Cavalli-Sforza, L.L., Majumder, P.P., and Underhill, P.A. Polarity and temporality of high-resolution Y-chromosome distributions in India identify both indigenous and exogenous expansions and reveal minor genetic influence of Central Asian Pastoralis. Amer. J. Human Genet. 78, 202221 (2006) The International HapMap Consortium. A second generation human haplotype map of over 3.1 million SNPs. Nature 449, 851-862 (2007). Weale, M.E., Yepiskoposyan, L., Jager, R.F., Hovhannisyan, N., Khudoyan, A. , Burbage-Hall, O., Bradman, N. and Thomas, M. Armenian Y chromosome haplotypes reveal strong regional structure within a single ethno-national group. Hum. Genet. 109, 659-674 (2001) Zalloua, P.A., Xue, Y., Khalife, J., Makhoul, N., Debiane, L., Platt, D.E., Royyuru, A.K., Herrera, R.J., Hernanz, D.F.S., Blue-Smith, J., Wells, R.S., Comas, D., Bertranpetit, J., Tyler-Smith et al. Y-Chromosomal diversity in Lebanon is structured by recent historical events. Amer. J. Hum. Genet. 82, 973-882 (2008). Zhivotovsky, L.A., Underhill, P.A., Cinnoglu, C., Kayser, M., Morar, B., Kivisild, T., Scozzari, R., Cruciani, F., Destro-Bisol, G., Spedini, G., Chambers, G.K., Herrera, R.J., Yong, K.K., Gresham, D., Tournev, I., Feldman, M.W., and Kalaydjieva, L. The effective mutation rate at Y chromosome short tandem repeats, with application to human population- divergence time. Am. J. Human Genet. 74, 50-61 (2004). |