- 1. Geographic distribution
- 2. Subclades
- 3. Origins
- 4. History & Subclades
- 4.1 Neolithic farmers & herders
- 4.2 Indo-European G2a3b
- 4.3 Roman redistribution
- 4.4 Scythian G1
- 5. MtDNA correspondance
- 6. Famous G2a individuals
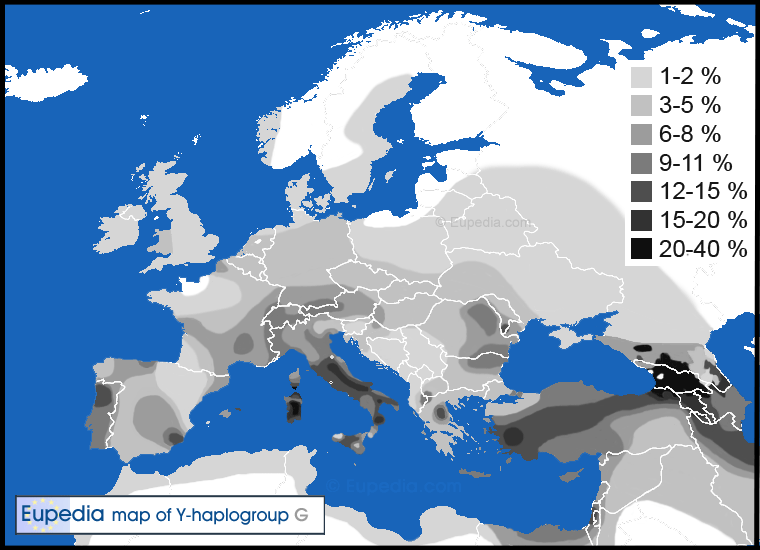
Geographic distribution
Nowadays haplogroup G is found all the way from Western Europe and Northwest Africa to Central Asia, India and East Africa, although everywhere at low frequencies (generally between 1 and 10% of the population). The only exceptions are the Caucasus region, central and southern Italy and Sardinia, where frequencies typically range from 15% to 30% of male lineages.Most Europeans belong to the G2a subclade, and most northern and Western Europeans more specifically to G2a-L141.1(or to a lower extend G2a-M406). About all G2b (L72+, formerly G2c) Europeans are Ashkenazi Jews. G2b has also been found around Afghanistan, probably as an offshoot of Neolithic farmers from the Levant.
Haplogroup G1 is found predominantly in Iran, but is also found in the Levant, among Ashkenazi Jews, and Central Asia (notably in Kazakhstan).
G2a makes up 5 to 10% of the population of Mediterranean Europe, but is fairly rare in Northern Europe. The only places where haplogroup G2 exceeds 10% of the population in Europe are Cantabria, central and southern Italy (esp. in the Apennines), Sardinia, northern Greece (Thessaly) and Crete - all mountainous and relatively isolated regions. Other regions with frequencies approaching the 10% include Asturias in northern Spain, Auvergne in central France, Switzerland, Sicily, the Aegean Islands, and Cyprus.
Distribution of haplogroup G in Europe, North Africa and the Middle East
Subclades

Origins
Haplogroup G descends from macro-haplogroup F, which is thought to represent the second major migration of Homo sapiens out of Africa, at least 60,000 years ago. While the earlier migration of haplogroups C and D had followed the coasts of South Asia as far as Oceania and the Far East, haplogroup F penetrated through the Arabian peninsula and settled in the Middle East. Its main branch, macro-haplogroup IJK would become the ancestor of 80% of modern Eurasian people. Haplogroup G had a slow start, evolving in apparent isolation for tens of thousands of years, possibly in Southwest Asia, cut off from the wave of colonisation of Eurasia.As of early 2014, there were 286 mutations (SNPs) defining haplogroup G, confirming that this paternal lineage experienced a severe bottleneck before splitting into happlogroups G1 and G2. Haplogroup G1 might have originated around modern Iran, while G2 would have developed in Southwest Asia during the Upper Paleolithic, probably in the Late Glacial period (19,000 to 12,000 years ago). At that time humans would all have been hunter-gatherers, and in most cases living in small nomadic or semi-nomadic tribes. Members of haplogroup G2 appear to have been closely linked to the development of early agriculture in the Levant part of the Fertile Crescent, starting 11,500 years before present. The G2a branch expanded to Anatolia, the Caucasus and Europe, while G2b ended up secluded in the southern Levant and is now found mostly among Jewish people.
There has so far been ancient Y-DNA analysis from only four Neolithic cultures (LBK in Germany, Remedello in Italy and Cardium Pottery in south-west France and Spain), and all sites yielded G2a individuals, which is the strongest evidence at present that farming originated with and was disseminated by members of haplogroup G (although probably in collaboration with other haplogroups such as E1b1b, J, R1b and T).
The highest genetic diversity within haplogroup G is found between the Levant and the Caucasus, in the Fertile Crescent, which is a good indicator of its region of origin. It is thought that early Neolithic farmers expanded from the Levant and Mesopotamia westwards to Anatolia and Europe, eastwards to South Asia, and southwards to the Arabian peninsula and North and East Africa. The domestication of goats and cows first took place in the mountainous region of eastern Anatolia, including the Caucasus and Zagros. This is probably where the roots of haplogroup G2a (and perhaps of all haplogroup G) are to be found. So far, the only G2a people negative for subclades downstream of P15 or L149.1 were found exclusively in the South Caucasus region.
History of haplogroup G
Several historical migrations brought different subclades of haplogroup G to Europe or redistributed them geographically.Neolithic farmers and mountain herders
The geographic continuity of G2a from Anatolia to Thessaly to the Italian peninsula, Sardinia, south-central France and Iberia already suggested that G2a could be connected to the Printed-Cardium Pottery culture (5000-1500 BCE). Ancient DNA tests were conducted on skeletons from numerous Neolithic European cultures and all yielded a majority of G2a samples. Remains have been tested to date for the Starčevo culture (6000-4500 BCE) Linear Pottery culture (5500-4500 BCE) in Hungary and Germany, as well as Cardium Pottery culture in southern France and north-east Spain. Nevertheless, substantial minorities of other haplogroups have been found on different Neolithic sites next to a G2a majority, including C1a2, F, I*, I1, I2a and I2a1 in Central and Southeast Europe (LBK, Starčevo) and E-V13 and I2a in the West Mediterranean (Cardium Pottery). These probably represent assimilated hunter-gatherers descended from Mesolithic and Paleolithic Europeans. It is interesting to note that many of those assimilated lineages, like C1a2, F and I*, are virtually extinct nowadays. Could it be that these people were ot assimilated after all, but rather enslaved by G2a tribes ?
Ötzi the Iceman (see famous individuals below), who lived in the Italian Alps during the Chalcolithic, belonged to haplogroup G2a2a2 (L91), a relatively rare subclade found nowadays in the Middle East, southern Europe (especially Sicily, Sardinia and Corsica) and North Africa. G2a2 (PF3146) is otherwise found at low frequencies all the way from the Levant to Western Europe. In conclusion, Neolithic farmers in Europe would have belonged to G2a, G2a2 (+ subclades) and G2a3 (and at least the M406 subclade).
Nowadays G2a is found mostly in mountainous regions of Europe, for example, in the Apennine mountains (15 to 25%) and Sardinia (12%) in Italy, Cantabria (10%) and Asturias (8%) in northern Spain, Austria (8%), Auvergne (8%) and Provence (7%) in south-east France, Switzerland (7.5%), the mountainous parts of Bohemia (5 to 10%), Romania (6.5%) and Greece (6.5%). It may be because Caucasian farmers sought hilly terrain similar to their original homeland, perhaps well suited to the raising of goats. But it is more likely that G2a farmers escaped from Bronze-Age invaders, such as the Indo-Europeans and found shelter into the mountains. For example, G2a3a (M406) is found at relatively high frequencies in the southern Balkans, the Apennines and the Alps, in contrast with G2a3b (L141.1), which is found everywhere in Europe.p>G2a people may have been among the first humans to have acquired the alleles for fair skin. A hunter-gatherer from northern Spain tested by Olalde et al. 2014 still had dark skinned as recently as 7000 years ago. In contrast, an Early Neolithic farmer from Germany possessed the alleles for fair skin found in modern Europeans. The Neolithic individual was female, but Neolithic men from the same LBK culture were predominantly G2a (but also included around 50% of assimilated Paleolithic/Mesolithic lineages such as C1a2, F, I1 and I2a). It is still unclear exactly when and among which haplogroup fair skin arose, but it has been suggested that the new diet brought by cereral agriculture would have caused deficiencies in vitamin D, which was traditionally absorbed from fish and meat among foragers. Mutations for light skin would have been positively selected among Neolithic agriculturalists to stimulate the production of vitamin D from sunlight in order to compensate for the scarcity of meat.
Expansion of agriculture from the Middle East to Europe (9500-3800 BCE)

G2a-L141.1, the Indo-European branch of G2a
Contrarily to other branches of G2a, which are more prevalent in mountainous areas, G2a3b (L141.1), and particularly the G2a3b1 (P303) subclade, is found uniformly throughout Europe, even in Scandinavia and Russia. More importantly, G2a3b and its subclades are also found in eastern Anatolia, the Caucasus, Central Asia and throughout India, especially among the upper castes, who represent the descendants of the Bronze Age Indo-European invaders. The combined presence of G2a3b1 across Europe and India is a very strong argument in favour of an Indo-European origin. The coalescence age of G2a3b1 also matches the time of the Indo-European expansion during the Bronze Age.
The homeland of R1b1a (P297) and Pre-Proto-Indo-European speakers is presumed to have been situated in eastern Anatolia and/or the North Caucasus. The Caucasus itself is a hotspot of haplogroup G. Therefore, it is entirely conceivable that a minority of Caucasian men belonging to haplogroup G (and perhaps also J2b) integrated the R1b community that crossed the Caucasus and established themselves on the northern and eastern shores of the Black Sea sometime between 7,000 and 4,500 BCE.
An alternative theory is that G2a3 (L30) came from Anatolia to eastern and Central Europe during the Neolithic (a fact proven by ancient DNA test). Once in Southeast Europe it split in two branches: G2a3a, who followed the Danube to Central Europe (LBK), and G2a3b, who migrated east to the Pontic Steppe and brought agriculture to the region. G2a3b would have mixed with the indigenous R1a people, then with R1b newcomers during the Chalcolithic and Bronze Age. By the time the Proto-Indo-Europeans started their massive expansion, G2a3b men (who apparently belonged overwhelmingly to G2a3b1 and its subclades) would have joined R1b-M269/L23 in the invasion of Old Europe from 4200 BCE (=> see R1b history). G2a3a would have been among the conquered populations of Old Europe, seeking refuge in mountainous areas.
Roman redistribution
By the Iron Age, the G2a population in most of Europe had been decimated by the Indo-European invasions, followed by Celtic warfare. G2a sought refuge from the invaders in the mountains, and like today, reached maximum frequencies in Italy (Apennines, Sardinia) and in the Alps.
The ancient Latins and Romans descend from the Italic tribes who invaded the Italian peninsula from 1200 BCE. They seem to have belonged primarily to haplogroup R1b-U152 (=> see Genetics of the Italian people), but to have carried a substantial minority of G2a3b (L141.1) lineages, especially the U1 and L497 subclades. The Latin homeland in central Italy is one of the hotspots for haplogroup G2a in Europe today. The high level in G2a in the Latium might be due to the dual presence of Indo-European G2a3b and of earlier Neolithic lineages who descended from the Apennines to live in Rome after being absorbed by the Roman civilisation.
If the ancient Romans and other Romanised peoples from the Italian peninsula had any genetic impact on other parts of the Roman Empire (as they should have), they certainly contributed to a moderate increase of G2a lineages (in addition to R1b-U152 and J2) within the borders of the empire. Indeed, the frequency of haplogroup G decreases with the distance from the boundaries of the empire. Haplogroup G is extremely rare Nordic and Baltic countries nowadays, despite the fact that agriculture reached those regions around the same time as Britain or Ireland. Another reason could be that the forested lowlands of northern Germany, Poland and the Baltic were too poor in metals and did not have attract as many Bronze-Age workers from the Caucasus (=> see Metal-mining and stockbreeding explain R1b dominance in Atlantic fringe). Northeast Europe also has a relatively low percentage of haplogroup R1b, which further reinforces the hypothesis that the two haplogroups spread together during the Bronze Age.
Scythian G1
Haplogroup G1 is the South and Central Asian branch of haplogroup G. While G2a men migrated west to Anatolia and Europe in the Neolithic, their G1 cousins migrated east to Persia and India. Only very rare cases of G1 have been found in Europe, including in Britain, Germany, as well as most of southern, central and eastern Europe. How did these G1 lineages get there ?
Central Asia became a merging zone for southern G1 and J2 lineages with northern R1a lineages during the Bronze and Iron Ages. New hybrid peoples were formed, like the Scythians, who once controlled an empire ranging from northern Pakistan to Xinjiang and to Ukraine. The Romans were known to recruit Scythian or Sarmatian horsemen in their legions. According to C. Scott Littleton in his book From Scythia to Camelot, several Knights of the Round Table were of Scythian origin, and the the legend of Holy Grail itself originated in ancient Scythia. This hypothesis was also taken up in the 2004 movie King Arthur, which opens with the arrival of Scytho-Roman cavalry in Britain. However, Scythians were steppe people more likely to belong to haplogroup R1a. If any of them did belong to G, they presumably were G1, not G2a. This would explain the scattered cases of G1 in north-Western Europe though.