В академической литературе за последние четверть
века регулярно появляются утверждения о том, что африканские ДНК-линии
«самые древние», хотя основания для того не приводятся. И понятно,
почему не приводятся – их просто нет. Давайте для начала разберемся, что
такое «древние ДНК-линии», понимая под этим цепочки снипов Y-хромосомы.

Но разбираться особенно и не надо, потому что у всех мужчин на Земле, африканцев и неафриканцев, во всяком случае, у всех, кто делал тест на ДНК, снип-цепочки Y-хромосомы по древности одинаковы. У всех эти цепочки снипов тянутся от общих предков с шимпанзе, орангутангом, гориллой, макакой и другими древними общими с нами предками. Ниже – характерный пример, процитированный еще в 2012 году в журнале Advances in Anthropology (Klyosov, Rozhanskii, Ryabchenko):

Сравнение фрагмента нуклеотидной последовательности Y-хромосомы
человека, шимпанзе, гориллы, орангутанга и макаки
Видно, что в 90% случаев нуклеотидные последовательности показанных фагментов Y-хромосомы совпадают. А у шимпанзе и человека совпадают 93 нуклеотида из 97, то есть в 96%. Так что у всех нас ДНК-корни древние, и африканцы в этом отношении никак не выделяются.
Хорошо, а насколько древние? Антропологи оценивают время расхождения линии будущего человека от линии будущего шимпанзе в 4-6 миллионов лет назад, это же повторяют и генетики, хотя поиски статей с конкретными расчетами результатов не дали. А вот результаты расчета с помощью автоматического калькулятора Килина-Клёсова (ККК, 2015), иллюстрация увеличивается по клику:

Общий предок человека и шимпанзе жил, если округлить расчеты, 4.2±1.5 миллионов лет назад. 16-маркерный гаплотип шимпанзе был извлечен из геномной базы данных, как описано в той статье в Advances in Anthropology (2012), а в качестве обобщенного гаплотипа человека брался экстраполированный предковый гаплотип по всем неафриканским гаплогруппам, пересчитанный на времена 100 тысяч лет назад. Впрочем, можно было и не пересчитывать, вот – время до общего предка современных носителей гаплогруппы А00 и шимпанзе, иллюстрация увеличивается по клику:

Оно равно 4.4±1.4 миллионов лет назад, что практически одинаково в пределах погрешности расчетов. Те же времена дают все основные гаплогруппы серии А, например:
• А0а-V150 имеет общего предка с шимпанзе 4.6±1.7 млн лет назад;
• A1b1-M32 (Чад и Турция) 4.3±1.4 млн лет назад;
• A1b1-M32, M13 (Британия) 4.2±1.4 млн лет назад, а также
• гаплогруппа В, 4.4±1.4 млн лет назад.
Короче, в отношении «древних ДНК-линий» африканцы ничем от неафриканцев не отличаются. И те, и другие тянут свои ДНК-линии от общего предка с шимпанзе. И у тех, и у других цепочки снипов уходят на глубины в сотни тысяч и миллионы лет. К этому мы еще вернемся.
Может, под «древними африканскими ДНК-линиями» попгенетики и прочие генетики подразумевают древние времена жизни их общих предков, и полагают, что они намного более древние, чем у неафриканцев? Придется их огорчить, ситуация противоположная. Африканские предки группы А (популяций, субкладов) почти все совсем недавние. Возьмем, например, гаплогруппу А00, которая среди гаплогрупп считается самой древней. Да, для нее цепочка снипов насчитывает – по данным, приводимым YFull – 1684 снипов, что при 144 годах на снип (это число приводит YFull) дает 242500 лет от начала гаплогруппы. Действительно, YFull приводят близкий «возраст» А00, 235 тысяч лет. Но откуда эта цепочка снипов ведет – никто не знает. Может, из Африки, может, с Кавказа, может, из Сибири, может, из Европы – неизвестно. Почему-то считается, что именно из Африки. Но это опять «по понятиям», правда, откуда появившимся, непонятно, извините за игру слов. Древнейшие ДНК находили в Европе, на Русской равнине, в Сибири, а в Африке ни разу, правда, и исследований таких не было. Так откуда эти «понятия» появились? А так, стиль попгенетиков.
Кстати, о возрасте гаплогруппы А00. Калькулятор КК дает для базовых гаплотипов А0 и А00 возраст общего предка (с округлением) 217000±87000 лет. Это – возраст А00 или немного старше. Так что датировка по снипам воспроизводится по гаплотипам в пределах погрешности расчетов (иллюстрация ниже увеличивается по клику).

Так вот, об общих предках африканских популяций, субкладов группы А. Они почти все по времени очень мелкие, часто всего несколько сотен лет. Иногда – между 1000 и 2000 лет. Самые древние – примерно 5000-7000 лет. И опять – где эти предки жили, никто не знает. Взглянем на гаплогруппу А00. В литературе (Mendez et al., 2013) описаны 11 гаплотипов в 95-маркерном формате (без 16 мультимаркерных гаплотипов) носителей гаплогруппы А00, все из племени Mbo из Камеруна, и еще один – афроамериканец. Расчетов в статье, конечно, нет. Все 11 гаплотипов содержат 29 мутаций, что дает 29/11/0.161 = 16 условных поколений, но с учетом того, что 45 аллелей (из 1045 аллелей) авторы определить не смогли, то до общего предка на самом деле 17 условных поколений, то есть 425±90 лет.
Итак, сама гаплогруппа А00 образовалась более 200 тысяч лет назад, причем неизвестно где, а общий предок носителей А00 из Камеруна жил примерно 425 лет назад. Если добавить к серии из Камеруна еще гаплотип афроамериканца той же гаплогруппы, который отличается от предкового гаплотипа камерунцев на 11 мутаций, то есть на 11/0.161 = 68 → 73 условных поколения, то есть на 1825 лет, то их общий предок, камерунцев и афроамериканца, жил примерно (425+1825)/2 = 1125 лет назад.
Примеров недавних общих предков разных мужских популяций гаплогрупп класса А, которые сейчас живут в Африке, можно привести немало. В большинстве этих популяций гаплотипы или одинаковы или почти одинаковы, даже для протяженных гаплотипов. С доступными гаплотипами гаплогруппы А00 мы уже разобрались, им четыре сотни лет. Взглянем теперь на Проект FTDNA гаплогруппы А:
• В гаплогруппе/субкладе А0-V150 имеются два 25-маркерных гаплотипа, оба идентичны друг другу, то есть их общий предок жил не более 300-400 лет назад. Других гаплотипов в 25-маркерном формате или выше нет.
• В гаплогруппе A0a-L981 (L994, V148, V166) между двумя доступными 67-маркерными гаплотипами есть 8 мутаций, что помещает их общего предка на 900 лет назад.
• В гаплогруппе A0b-V166, L896 шесть 37-маркерных гаплотипов имеют общего предка 475±160 лет назад.
• В гаплогруппе A1a-M31 («европейский кластер») пять 37-маркерных гаплотипов имеют общего предка 1175±290 лет назад.
• В гаплогруппе A1a-M31 (все неизвестного происхождения) пять 37-маркерных гаплотипов имеют общего предка 1880±390 лет назад.
• В гаплогруппе A1a-M31 (из 18 гаплотипов – 14 неизвестного происхождения, два из Англии, один из Швейцарии, один Кейп Верде) все 37-маркерные, все имеют общего предка 730±130 лет назад.
• В гаплогруппе A1a-M31 («британский кластер») 19 гаплотипов в 67-маркерном формате имеют общего предка 740±120 лет назад.
В итоге, 50 гаплотипов в 37-маркерном формате четырех предшествующих серий (А0а-М31) имеют общего предка 9270±970 лет назад. Но надо обратить внимание, что это четыре серии гаплотипов, разбросанные по разным континентам. Это – не одна «ДНК-линия», это – четыре разных ДНК-линии, с четырьмя общими предками, каждый «глубиной» от 700 до 1900 лет.
Ну и где, позволительно спросить, «древнейшие ДНК-линии в Африке»? Это – миф, ничем не обоснованный. Ископаемых ДНК нет, древних линий, как мы видим, нет, к тому же, в Африке вовсе не гаплотипы группы А явяются доминирующими. Доминирует гаплогруппа E1b; в Камеруне и Чаде, и отчасти в Нигерии, доминирует гаплогруппа R1b, поэтому неудивительно, что в Африке наблюдается «высокое генетическое разнообразие», только это не от древности, а от обилия носителей разных гаплогрупп, прибывших в Африку в разные времена. Да и вообще, как можно серьезно обращать внимание на «генетическое разнобразие» в открытых системах, какой является Африка? Как я уже не раз отмечал, самым «генетически разнообразным» городом мира является, наверное, Нью-Йорк, но вряд ли кто будет серьезно утверждать, что Нью-Йорк – древнейший город мира. Кроме, наверное, популяционных генетиков, с их примитивными и бездумными критериями.
Откуда же появился этот миф о «древности африканских линий»? Датировок ведь их до последнего времени не было, да и датировки бывают, как мы убедились, нескольких видов – цепочек мутаций, уходящих далеко вглубь, до приматов; цепочек мутаций, ведущих ко временам образования соответствующих гаплогрупп; и датировок общих предков современных носителей соответствующих гаплогрупп и субкладов. Ни одна из этих датировок не «укоренена» территориально, то есть мы не знаем, на какой территории, на каком континенте образовалась та или иная гаплогруппа. Мы можем только набирать сведения и складывать фрагменты общей картины, но Африка в этом отношении находится на одном из последних мест по накопленным знаниям. Но фантазий, натяжек и откровенных фальсификаций про них со стороны попгенетиков – хоть отбавляй.
Собственно, это характерно для популяционных генетиков, но они уроки из принципиальных ошибок не извлекают. Еще не так давно они утверждали, что гаплогруппа R1b образовалась в Европе 30 тысяч лет назад, и сначала приняли в штыки, а потом напрочь игнорировали сведения и датировки ДНК-генеалогии, что R1b прибыла из Южной Сибири длинным миграционным путем в Европу примерно 4800 лет назад (Klyosov, 2009; Klyosov, 2012). Называли это «псевдонаукой», особенно в этом усердствовали попгенетики Балановские. Разворот произошел в 2013 году, когда неподалеку от Байкала нашли ископаемую гаплогруппу R с археологической датировкой 24 тысячи лет назад. Лед тронулся, и попгенетики, не моргнув глазом, не повинившись и не цитируя статьи с датировками ДНК-генеалогии, стали повторять то, что уже было опубликовано нами несколько лет назад. Например, в недавней статье Batini, Jobling и других (2015) датировки гаплогруппы R1b в Европе уже даются как R1b-L11 4510 лет (доверительный интервал 3920-5160 лет назад) и R1b-P312 4210 лет (доверительный интервал 3700-4790 лет назад). YFull дают датировки R1b-U106 4900 лет (доверительный интервал 4400-5400 лет назад), R1b-P312 4600 лет (для TMRCA, доверительный интервал 4300-5000 лет назад).
К сожалению, приходится повторять, что попгенетики уроков из своих ошибок не извлекают. Ситуация повторяется с Африкой. Данных нет, или данные говорят о другом, но их придумывают и искажают. Зачем это им нужно? Для нормального человека звучит невероятно, но это ментальность вассалов, которые лебезят перед сюзереном. Сюзерен решает, кого пригреть, кого пригласить на конференцию, кому дать грант, кого взять в коллективную статью, ввести в комиссию или комитет, в общем, в инфраструктуру. Без сюзерена вассал – никто, и вассал это прекрасно понимает. А зачем это нужно сюзерену? О, там большая политика. Вассала к этим вопросам не допускают. Вассал знает, что если он публично усомнится в африканском происхождении человечества, то его дни сочтены. Поэтому он, вассал, готов разорвать на части любого, кто сомневается или выставляет данные против. А поскольку он разорвать на части не может, то цель – найти компромат, состряпать ложь, передернуть, исказить – других способов или аргументов у вассала нет, во всяком случае, научных.
Посмотрим на схему гаплогрупп и субкладов группы А, приведенную согласно классификации ISOGG, в сокращенном виде:

В этой схеме – источник принципиальной путаницы попенетиков. Они не смогли увидеть и осознать, что эта лесенка субкладов представляет собой серию вилок, которые периодически расходятся на «африканские» (по сегодняшнему месту жительства) и неафриканские ветви. Опишем кратко эти вилки, а потом рассмотрим их подробнее – когда они образовались, и кто являются потомками субкладов по обе стороны тех вилок.
Итак, вилок всего четыре. Первой от ствола, уходящего вглубь эволюционного дерева человечества (точнее, его Y-хромосомы), отходит вилка, знаменующая, видимо, начало Homo sapiens – расхождение на А00 и А0-Т. Вторая – расхождение А0-Т на А0 и А1. Третья – расхождение А1 на А1a и A1b. Четвертая – расхождение A1b на A1b1 и ВТ.
Если упрощенно – то каждая вилка расходится на африканскую (по нынешнему месту жительства) и неафриканскую стороны. А00, А0, A1a и A1b1 уходят в Африку, противоположные стороны вилки A0-T, A1, A1b и BT остаются вне Африки. «Упрощенно» – потому что вилка на А00 и A0-T образовалась еще до расхождения на африканцев и неафриканцев (это расхождение произошло примерно 160 тысяч лет назад), а впоследствии многие неафриканцы перебирались в Африку – это и многочисленные носители гаплогрупп E1b, J2, R1b, и так далее. Среди них – миллионы современных жителей Западной Африки (берберы, жители Камеруна и Чада), египтяне, потомки буров в Южной Африке, и так далее. Немало жителей Европы и Ближнего Востока имеют гаплогруппу A1a, как недавно показывал И.Л. Рожанский в статье на Переформате. Да и вообще носителей группы А в Африке относительно мало – часто на уровне единиц процента или того меньше. Рассмотрим вилки подробнее.
Первая вилка – расхождение предшественника Homo sapiens (или Homo sapiens в начальной стадии, примерно 230 тысяч лет назад) на гаплогруппы А00 и А0-Т. Нисходящих субкладов от А00 пока не обнаружено, и ее нынешние носители обнаружены только в Африке в количестве 11 человек (см. выше), с общим предком несколько сотен лет назад, а также несколько афроамериканцев с одинаковыми гаплотипами. Снипы гаплогруппы А0-Т, напротив, имеются у всех неафриканцев, в том числе у всех нас, читателей Переформата, хотя только у мужчин. И, разумеется, у всех африканцев, носителей гаплогрупп E1b, J2, R1b, В и других, которые прибыли в Африку относительно недавно. Но помимо них снипы группы А0-Т должны быть и у всех африканцев, носителей групп А, потому эти снипы предшествуют разделению африканских и неафриканских линий (примерно 160 тысяч лет назад). Впрочем, речь идет о промежуточных снипах в цепочках наших и африканских снипов, носителей А0-Т как терминальных снипов пока ни у кого не выявлено, ни в Африке, ни вне Африки. Поэтому называть его «африканским» никакого смысла не имеет.
К тому, что у всех нас, мужчин, есть множество снипов гаплогруппы А0-Т – ниже приведен список части из 587 снипов этой гаплогруппы, из которых у меня есть L1090, L1121, L1124, L1125, L1127, L1235 и многие другие:
Мой список снипов BigY
CTS10042, CTS10627, CTS10834, CTS10847, CTS10976, CTS10993, CTS11148, CTS11184, CTS11411, CTS11468, CTS11530, CTS1169, CTS11720, CTS11734, CTS11853, CTS11913, CTS11948, CTS12010, CTS1211, CTS1277, CTS1278, CTS12947, CTS12972, CTS1340, CTS1367, CTS1415, CTS1416, CTS1417, CTS1419, CTS1422, CTS1619, CTS1738, CTS1795, CTS1907, CTS2134, CTS262, CTS2626, CTS2636, CTS2637, CTS2638, CTS2639, CTS2663, CTS 2680, CTS2891, CTS2988, CTS2992, CTS3067, CTS3075, CTS3229, CTS3230, CTS3231, CTS3316, CTS3321, CTS3358, CTS3402, CTS3441, CTS3527, CTS3534, CTS3548, CTS3551, CTS358, CTS3607, CTS3622, CTS3649, CTS3650, CTS3654, CTS3697, CTS3775, CTS3848, CTS3943, CTS3984, CTS4259, CTS4465, CTS4862, CTS501, CTS5287, CTS5423, CTS5454, CTS5508, CTS5577, CTS5648, CTS570, CTS5936, CTS5979, CTS623, CTS6918, CTS6985, CTS7191, CTS7275, CTS7278, CTS7400, CTS7500, CTS7650, CTS7659, CTS7941, CTS8008, CTS8127, CTS8350, CTS8507, CTS8508, CTS8591, CTS8626, CTS8665, CTS8797, CTS8816, CTS8896, CTS9018, CTS903, CTS9108, CTS9510, CTS9515, CTS9539, CTS9596, CTS9733, CTS9739, CTS9754, CTS9779, F1050, F1209, F1224, F132, F1586, F184, F185, F2957, F3032, F313, F3159, F3166, F3185, F3194, F3197, F3337, F3561, F3564, F3644, F3650, F3692, F4099, F686, F719, F773, F82, F937, F989, IMS-JST002612, IMS-ST022448_3, IMS-ST022448_6, IMS-ST022449_3, IMS-ST022449_6, IMS-ST022451_2, K511, L1002, L1004, L1005, L1009, L1013, L1028, L104, L105, L1053, L1060, L1061, L1062, L1067, L1071, L108, L1089, L1090, L1095, L1098, L1099, L110, L1101, L1105, L1112, L1116, L1118, L112, L1120, L1121, L1123, L1124, L1125, L1127, L1129, L113, L1132, L1135, L1136, L1137, L114, L1142, L1143, L1145, L1150, L1155, L1179, L1185, L1120, L1225, L1235, L132, L1346, L1347, L1352, L1480, L1492, L15, L16, L168, L248, L268, L313, L350, L352, L413, L418, L438, L440, L449, L457, L468, L469, L498, L543, L58, L604, L721, L74, L741, L747, L76, L760, L768, L777, L779, L781, L819, L82, L821, L851, L875, L882, L962, L969, L970, L977, L986, L989, M168, M173, M198, M207, M213, M235, M251, M294, M299, M306, M417, M42, M420, M448, M449, M45, M459, M511, M512, M513, M514, M515, M516, M526, M74, M89, M9, M94, P108, P128, P131, P132, P133, P134, P135, P136, P138, P139, P14, P140, P141, P143, P145, P146, P148, P149, P151, P157, P158, P159, P160, P161, P163, P166, P187, P19_1, P207, P224, P225, P226, P228, P229, P230, P231, P232, P233, P234, P235, P236, P237, P238, P239, P242, P243, P244, P245, P27.1_2, P280, P281, P282, P283, P284, P285, P286, P294, P305, P316, PAGES00026, PAGES00081, PAGES00083, PF1345, PF1378, PF1389, PF1396, PF1402, PF1403, PF1405, PF1407, PF1408, PF1411, PF1412, PF1414, PF1415, PF1418, PF1419, PF2589, PF2590, PF2591, PF2608, PF2610, PF2611, PF2614, PF2615, PF2616, PF2619, PF2620, PF2621, PF2622, PF2626, PF2629, PF2631, PF2634, PF2635, PF2637, PF2640, PF2642, PF2647, PF2651, PF2653, PF2658, PF2660, PF2683, PF2685, PF288, PF2690, PF2694, PF2695, PF2700, PF2709, PF2716, PF2718, PF2722, PF2723, PF2729, PF2731, PF2732, PF2736, PF2737, PF2739, PF2742, PF2747, PF2748, PF2749, PF2752, PF2753, PF2754, PF2755, PF2756, PF2757, PF2758, PF2770, PF2772, PF2988, PF349, PF3494, PF3495, PF3496, PF3497, PF3500, PF4522, PF5459, PF5469, PF5477, PF5483, PF5488, PF5490, PF5498, PF5500, PF5503, PF5513, PF5861, PF5862, PF5863, PF5864, PF5869, PF5870, PF5871, PF5872, PF5876, PF5881, PF5882, PF5885, PF5886, PF5888, PF5889, PF5895, PF5908, PF5911, PF5914, PF5916, PF5919, PF5920, PF5936, PF5940, PF5943, PF5944, PF5945, PF5949, PF5951, PF5953, PF5954, PF5955, PF5956, PF5957, PF5958, PF5964, PF5965, PF5967, PF5970, PF5971, PF5975, PF5978, PF5980, PF5981, PF5982, PF5984, PF5985, PF5991, PF6013, PF6016, PF6047, PF6054, PF6056, PF6057, PF6066, PF6069, PF6071, PF6072, PF6073, PF6114, PF6116, PF6118, PF6121, PF6132, PF6133, PF6143, PF6153, PF6156, PF6158, PF6159, PF6160, PF6162, PF6163, PF6165, PF6168, PF6169, PF6170, PF6172, PF6179, PF6181, PF6194, PF6195, PF6201, PF6204, PF6211, PF6212, PF6213, PF6214, PF6215, PF6216, PF6217, PF6218, PF6220, PF6222, PF6230, PF6231, PF6233, PF6512, PF682, PF7514, PF7522, PF7527, PF7530, PF7531, PF7532, PF7534, PF7540, PF7542, PK1, PR1220, PR202, PR2119, PR5761, PR855, rs161492_1, V221, V238, V241, V250, Y2902, Y32, Y33, Y34, Y36, Y783.2, YSC0000057, YSC0000077, YSC0000081, YSC0000107, YSC0000108, YSC0000109, YSC0000110, YSC0000193, YSC0000350, YSC0000913, Z1331, Z280, Z282, Z5997, Z6030, Z6032.
Фрагмент из списка снипов гаплогруппы A0-T (приведены 182 снипа из 587)

Понятие «вилка» не стоит понимать буквально, А00 и А0-Т могли образоваться с разницей в десятки тысяч лет. По данным YFull, до начала А00 насчитывается 1684 снипа, до начала А0-Т – 587 снипов, но возраст обеих гаплогрупп почему-то выставлен как одинаковый, 235 тысяч лет назад. На такую повторяющуюся «дихотомию» в презентации данных YFull мы уже не раз обращали внимание. В любом случае, искать у нас, читателей Переформата, снипы гаплогруппы А00 бесполезно, нет их у нас, ни одного из тех 1684 снипов. Но важно то, что А0-Т образует очередную вилку, на гаплогруппы А0 и А1.
Вторая вилка – расхождение А0-Т на А0 и А1. Потомки А0 сейчас живут в Африке, потомки А1 – мы с вами, читатели Переформата. У всех нас в цепочках снипов есть снипы субклада А1 (см. ниже), но это опять не значит, что наши предки вышли из Африки. Это опять промежуточные снипы, и никого с терминальными снипами А1 пока не выявлено, ни в Африке, ни вне Африки. Поэтому называть его «африканским» никакого смысла тоже не имеет.
По данным, приведенным YFull, к A0 ведет 899 снипов (это 129460 лет, если снип образуется в среднем раз в 144 года, как считали те же YFull), к А1 – 62 снипа (8900 лет), возраст А0 не выставлен, но для А1 выставлено 146300 лет «возраста». Опять загадка, как это может быть.
Судя по этим данным, на протяжении 100 тысяч лет между А00 и А0-Т, гаплогрупп между ними вовсе не образовывалось, и к чему те 62 снипа относятся – непонятно. Но здесь важнее то, что А0 – это «африканская» гаплогруппа (опять по нынешнему месту жительства), а А1 – неафриканская. На схеме выше показано семь субкладов гаплогрупы А0, и снипов никаких из них у нас, читателей Переформата, нет, а снипы гаплогруппы А1 есть у нас всех (я не буду больше повторять, что это относится к обладателям Y-хромосомы, то есть к мужчинам, и что «читатель Переформата» относится к неафриканцам, потомкам гаплогруппы ВТ, которые не перебрались на жительство в Африку).
Повторю, что «африканских» снипов А0 у нас нет, можно не искать, нет ни одного из 899 тех снипов. Не выходили наши предки из Африки, или, во всяком случае, не являются потомками ни А00, ни А0.
Надо сказать, что в Африке носителей гаплогруппы А0 обнаружено немного. Scozzari et al (2012) указывали, что только после долгих поисков нашли трех (!) носителей гаплогруппы А0, все в Камеруне, и отмечали, что эта гаплогруппа имеет в Африке очень низкую частоту встречаемости.
Ниже приведен список из 62 снипов гаплогруппы А1, из которых у меня есть L986, L989, L1002, L1009, L1112, V238, V241, V250. Остальных нет потому, что не все снипы у каждого определяются.
Список снипов гаплогруппы A1

Третья вилка – расхождение А1 на A1a и A1b. Потомки A1a сейчас живут в Африке и по всему миру (в частности, в ряде европейских стран и на Ближнем Востоке). Снипы А1b есть у всех нас, читатели Переформата. Но это опять не значит, что наши предки вышли из Африки. Это опять промежуточные снипы, и никого с терминальными снипами А1b пока не выявлено, ни в Африке, ни вне Африки. Поэтому называть его «африканским» опять никакого смысла не имеет. По данным, приведенным YFull, к A1a ведет 495 снипов (это 71 тысяча лет, если снип образуется в среднем раз в 144 года), к А1b – 44 снипа (6300 лет), но почему-то им опять было выставлено по 132600 лет «возраста». Опять загадка, на каком основании так делается.
Хотя носители гаплогруппы A1a разбросаны по всему миру (поэтому в любом случае это не «африканская» гаплогруппа, даже по такому условному критерию, как современное место жительства), их мало. И.Р. Рожанский уже приводил в своей статье «Африканцы на севере Европы. Африканцы ли?», что гаплогруппа A1a-M31 чрезвычайно редкая, и даже в Африке ее наличие исчисляется в лучшем случае 4.4% (в Западной Африке – Гвинея-Бисау, Гамбия, Сенегал, Мали, Нигер, Марокко), и 0.4% среди афроамериканцев. Этому же вторит статья Scozzari et al. (2012), что им удалось найти в Африке для геномных иследований только двоих (!) носителей A1a-M31.
Ниже приведен список из 44 снипов гаплогруппы А1b, но в той системе, которую использует YFull. В классификации ISOGG в гаплогруппе A1b насчитывается 47 снипов, и ни один не пересекается по индексам с приведенными ниже. Из этих 47 снипов у меня есть L989, L1009, L1013, L1053, P108, V221. Остальных нет, потому что, как указывалось выше, не все снипы у каждого определяются.
Список снипов гаплогруппы A1b (согласно классификации, используемой YFull)

И вот здесь снипы гаплогруппы A1b приоткрывают загадку. Воспроизведем еще раз фрагмент лесенки снипов гаплогруппы A1:

Мы видим, что от нее вилкой отходят субклады A1a и A1b. Мы уже узнали, что А1а встречается в Европе, на Ближнем Востоке, в Африке, причем нет никаких данных, что он изначально был африканским. Субклад A1b встречается у всех неафриканцев как промежуточный, в цепочке снипов. Субкладов, нижестоящих от A1b, а это A1b1, A1b1a, A1b1a1, A1b1a1a, A1b1a1a1, A1b1a1a2, A1b1a1a2a, A1b1a1a2b – у неафриканцев нет, возможно, потому, что параллельно с A1b1 образовался субклад BT (по даным YFull примерно 126 тысяч лет назад), и A1b1 ушли в Африку, а ВТ остался вне Африки. То есть у неафриканцев имется основная линия A1b > BT, и потомки ее составляют все гаплогруппы от В до Т. И вдруг выясняется, что нижестоящая линия, в субкладе A1b1a1- M23, P3, обнаружилась в Южной Африке среди носителей койсанских языков. Но это – довольно далекий по цепочке вниз субклад, который, по-видимому, отошел от неафриканцев в древние времена. Иначе говоря, не неафриканцы отошли от африканцев, а наоборот, африканцы отошли от неафриканцев, мигрировав в Африку. Опять – не выходили наши предки из Африки. Было наоборот: современные африканцы – потомки неафриканцев.
Четвертая вилка – расхождение A1b на A1b1 и BT. A1b1 и нисходящие субклады считаются африканскими – по нынешнему месту проживания его носителей, хотя родительская гаплогруппа A1b (как промежуточный субклад в цепочке снипов) насчитывает не менее 2-3 миллиардов неафриканцев, а нижеследующие субклады от A1b1 и ниже – обнаружены всего у нескольких африканцев. Вот и считайте, кто от кого произошел. Ответ ясен – африканцы от неафриканцев. Повторю – не выходили наши предки из Африки.
Итак, потомки A1b1 сейчас живут в минимальном количестве в Африке, а потомки гаплогруппы ВТ – это все неафриканские гаплогруппы от В до Т. По данным YFull, к A1b1 ведут всего 26 снипов, к субкладу A1b1a ведут всего 2 снипа, к A1b1a1 – 11 снипов, ни одного из них у нас нет. Датировок ни к одному из нисходящих от A1b1 субкладов в списке YFull нет, и понятно, почему – их носители крайне редко встречаются, да и данных нет. Снипы ВТ есть у всех нас, читатели Переформата.
Список снипов гаплогруппы A1b1 (согласно классификации, используемой YFull)

К сводной гаплогруппе ВТ ведут 403 снипа, и многие из них есть у нас. У гаплогрупп категории А их, естественно, нет. Датировка ВТ по данным YFull – примерно 126 тысяч лет, хотя 403 снипа – это примерно 58 тысяч лет. Некоторые могут подумать, что эти снипы – от родительской гаплогруппы A1b, но та датирована той же компанией YFull примерно 133 тысячами лет назад, то есть всего на 7000 лет раньше. Опять остается неясным, к чему те 403 снипа относятся, и откуда тянутся. Самая загадка в том, как «параллельные» гаплогруппы A1b1 и BT, от одного общего предка A1b, и к которым идут 26 и 403 снипа, соответственно, могут оказаться с одинаковой датировкой – 126 тысяч лет.
Ниже приведен частичный список снипов сводной гаплогруппы ВТ. Как уже отмечалось, многие из них есть у каждого неафриканского мужчины. У меня есть, например, L418, L604, L962, L970, L977, PF1408 и другие.
Фрагмент из списка снипов гаплогруппы BT (приведены 166 снипов из 403)

Итак, первая часть этой статьи безоговорочно показывает, что «африканские ДНК-линии» совсем не древние, в противовес тому, чем нас пичкают популяционные генетики уже 30 лет. Если вести «древность» от общих предков с приматами, то африканские линии ничем не отличаются от неафриканских, имеют одну и ту же глубину. Если считать от времени жизни общих предков современных мужских популяций – по племенам или по территориям, то африканские ДНК-линии весьма мелкие, намного мельче, чем неафриканские. Если считать от образования гаплогрупп, то никто не знает, на каких территориях или континентах они образовались, поэтому присваивать им названия «африканские» – некорректно. Более того, носителей этих гаплогрупп, категории А, в Африке очень мало. К этому могут быть разные причины – прошли бутылочные горлышки популяции, вымерли, или прибыли в Африку относительно недавно – большого значения не имеет, они не «древние», в том смысле, который вкладывают попгенетики. В итоге имеем то, что изображено на заглавной иллюстрации поста.
Так что с этим разделом закончили – нет в Африке «древних ДНК-линий». Но есть такое понятие, как совокупные ДНК-линии. Это тогда, когда сами ДНК-линии мелкие, но они – ошметки древних раскидистых деревьев гаплогрупп и субкладов. Мы видим только верхушки, но экстраполяция верхушек, в данном случае гаплотипов этих «ошметок», позволяет увидеть, когда жили общие предки этих разбросанных гаплотипов. Мы уже видели это выше – сравнение гаплотипов «мелких» популяций гаплогрупп А00 и А0 показывает, что они расходятся на астрономические расстояния. Сами «ошметки» имеют общих предков всего несколько сотен лет назад, но они расходятся на полмилиона лет, и отсюда следует, что их общие предки жили около 250 тысяч лет назад. В этом отношении африканские ДНК-линии отличаются от неафриканских. Рассмотрим это подробнее, в этом упрятана одна из основных загадок человечества.
Загадка была впервые сформулирована в 2012 году, в статье (Klyosov, Rozhanskii, Advances in Anthropology, 2, No. 2, 80-86). Суть ее состояла в том, что все ДНК-линии современных гаплогрупп от В до Т, то есть неафриканских гаплогрупп, сходятся к общему предку, который жил 64000±6000 лет назад, хотя разрозненные линии африканских (по нынешнему месту жительства) гаплогрупп сходятся к общему предку 132 тысяч лет назад (хотя сами разрозненные линии датируются временами жизни общих предков всего лишь 5500, 5000 и 600 лет назад). При этом все неафриканские линии (включая гаплогруппу В) отстоят исключительно далеко от африканских линий, ясно показывая, что неафриканские линии не могли произойти от африканских. Например, базовые гаплотипы гаплогрупп А0 и В отстоят друг от друга на 270 тысяч лет (между ними 18 мутаций в «медленных» 22-маркерных гаплотипах; для сравнения, базовые гаплотипы R1a и R1b отстоят друг от друга всего на 8 мутаций, то есть примерно на 54 тысячи лет, и это помещает их общего предка на 27 тысяч лет назад), иллюстрация увеличивается по клику.

Ясно, что если между гаплогруппами А0 и В 270 тысяч лет, то вторая никак не могла образоваться от первой в течение всего периода существования Homo sapiens, не говоря о 70 тысячах лет назад, которые попгенетики отводят для времени образования гаплогруппы В из А. Это аналогично примеру выше для R1a и R1b, в котором 54 тысячи лет расстояния между ними никак не могут допустить, что одна образовалась из другой; они обе образовались от одного общего предка, которым в данном случае являлась гаплогруппа R1.
Гаплогруппа А0, далеко отстоящая от гаплогруппы В, здесь не исключение. Подобные огромные расстояния отделяют гаплогруппу В от любых гаплотипов в субкладах категории А, например, 250 тысяч лет от A0a-L981, 280 тысяч лет от A1b-M32, 310 тысяч лет от европейского кластера гаплогруппы A1a-M31, 360 тысяч лет от другого кластера той же гаплогруппы, 320 тысяч лет от арабского кластера той же гаплогруппы, 350 тысяч лет от A0a-V150, 356 тысяч лет от A0a-L979, 346 тысяч лет от A0a1a-V151, и так далее. Самое большое расстояние отделяет гаплогруппу В от А00 – 450 тысяч лет, то есть их общий предок жил примерно 225 тысяч лет назад, на утренней заре Homo sapiens.
Такая же картина наблюдается и для гаплотипов всех гаплогрупп от В до Т. Например, растояние от гаплогруппы С до А0-L896 составляет 357 тысяч лет, до A0a1-V150 – 300 тысяч лет, и так далее. Но расстояние от В до С составляет всего 120 тысяч лет, то есть их общий предок жил примерно 60 тысяч лет назад. Это находится в полном соответствии с картиной, что гаплогруппа В образует единый кластер с гаплогруппами от В до Т, и не является по происхождению «африканской» гаплогруппой.
Видимо, уже всем ясно, что гаплогруппа В не образовалась из какой-либо гаплогруппы категории А (поскольку нет гаплогруппы А как таковой, есть А00, А0-Т, А0, А1, А1a, A1b и так далее), а все они ведут свое начало от общего предка. Этот общий предок жил примерно 160 тысяч лет назад, как показано в упомянутой выше статье (Klyosov, Rozhanskii, Advances in Anthropology, 2, No. 2, 80-86). Если исходить из датировок, приведенных на сайте YFull, то основатели гаплогрупп А00 и A0-T жили раньше этого времени, причем гаплогруппа А00 осталась в известном смысле тупиковой, нисходящие от нее субклады и прочие ДНК-линии неизвестны. A0-T прошла своими снипами как к африканцам, так и к неафриканцам, как показано выше, поскольку образовалась еще до разделения на африканскую и неафриканскую ветви. Разделение на эти ветви соответствует временам непосредственно до образования гаплогрупп А0 и А1, причем А0 ушла в Африку, А1 осталась вне Африки. Впоследствии носители многих гаплогрупп мигрировали в Африку, например, A1a и нижестоящие субклады, A1b1 и нижестоящие субклады, поэтому их снипов у нас нет, как описано выше.
Все это описано в целом в упомянутой выше статье (Klyosov, Rozhanskii, Advances in Anthropology, 2012), которая, кстати, набрала рекордное количество просмотров (около 90 тысяч) и скачиваний (около 20 тысяч), но академическое сообщество предпочло ее не заметить. Слишком многое она меняет в представлениях. Основная иллюстрация в той статье следующая:

Номенклатура в этой иллюстрации несколько устаревшая, в ней нет гаплогруппы А00, которая в 2012 году была неизвестной, но суть ее остается правильной. Гаплогруппа альфа – это условная гаплогруппа, от которой пошло разделение эволюционного Y-хромосомного дерева человечества на африканскую (слева) и неафриканскую линии. В настоящее время все гаплогруппы от В до Т, то есть гаплогруппы неафриканского происхождения, сходятся к условной же бета-гаплогруппе, которой на самом деле является сводная гаплогруппа ВТ. Загадка – куда делась связка между альфа- и бета-гаплогруппой, протяженностью в сто тысяч лет? И, кстати, куда делись африканские ДНК-линии, которые на левой части диаграммы полностью экстраполированы, от общих предков всего от тысячи до нескольких тысяч лет давности, вглубь времен?
Со времени публикации той статьи прошло уже более трех лет, в базах данных и в Проектах с тех пор накопилось много гаплотипов, в том числе протяженных, 111-маркерных, и появилась возможность проверить количественные выводы статьи. То, что расхождение африканской и неафриканской ДНК-линий произошло примерно 160 тысяч лет назад, мы уже проверили. Перейдем ко времени жизни общего предка всех неафриканских гаплогрупп – от В до Т. Ниже – дерево гаплотипов этих гаплогрупп. Оно построено по 111-маркерным гаплотипам, и сами гаплогруппы показаны по окружности дерева. Для каждой из 18 гаплогрупп использовали 10-25 гаплотипов, взятых произвольно из выборки, всего 403 гаплотипа nd 111-маркерном формате. Гаплогруппы категории А на дереве не оставили, так как они явно «вылезали» из кругового дерева, поскольку, как мы уже показали, они удалены по мутациям от всех других гаплогрупп.

Видно, что все гаплогруппы от В до Т образуют достаточно единую систему ДНК-линий (выраженную здесь через 111-маркерные гаплотипы), так что про некую «африканскую гаплогруппу В» говорить не приходится. Она такая же «автохтонная африканская», как R1b – «автохтонная европейская». Это застарелая болезнь попгенетиков – то, что видят сейчас, значит, так было всегда. Болезнь называется «отсутствие научного подхода», или «отсутствие научной школы», что, впрочем, одно и то же, а если по-русски, то «думать не умеют».
403 гаплотипа в 111-маркерном формате, то есть почти 45 тысяч аллелей – это крепкий орешек, если считать вручную. Более того, за десятки тысяч лет в них накапливаются десятки тысяч мутаций, помимо того, огромное количество возвратных мутаций, что делает 111-маркерную систему такой древности неустойчивой, и неприемлемой для расчетов, поскольку датировки значительно занижаются. Поэтому для данных случаев разработана 22-маркерная панель из самых «медленных» маркеров. Они настолько устойчивы во времени, что для всех 403 гаплотипов за десятки тысяч лет маркер DYS472 вообще не мутировал ни разу, DYS436 – всего 30 раз, DYS590 – 42 раза, и так далее. Сравните – маркер DYS390 за это же время должен мутировать 2270 раз, то есть в среднем 5.6 раз на гаплотип. При таких частотах мутации расчет вклада обратных мутаций уже практически невозможен, система нестабильна.
А вот «медленная» 22-маркерная панель с малыми скоростями мутаций позволяет проводить расчеты на десятки и сотни тысяч лет вглубь, и даже на миллионы лет, как было показано выше на примере расчета времени жизни общих предков человека и шимпанзе. Более того, разработанный калькулятор Килина-Клёсова проводит расчеты по каждому маркеру отдельно, используя квадратичный метод, и поэтому не требует введения поправок на возвратные мутации. В итоге ДНК-генеалогия получила незаменимый метод расчета массивных серий гаплотипов в любом формате и любой древности общего предка (Килин и Клёсов, Вестник Академии ДНК-генеалогии, 2015, т. 8, № 3, 321-375, http://dna-academy.ru/kilin-klyosov/). Расчет дерева из 403 гаплотипов, приведенного выше, показал, что общий предок гаплогрупп от В до Т жил примерно 63 тысячи лет назад (результат без округления – 63036 лет, что, конечно, с такой точностью совершенно излишне). Это почти точно совпадает с рассчитанной ранее (Klyosov and Rozhanskii, 2012) величиной 64±6 тысяч лет.
Теперь осталось подойти к решению сформулированной в той статье загадки – повторим ее опять: куда же делись предки Y-хромосомных линий в Африке ранее 5-10 тысяч лет назад, и у неафриканцев ранее 64±6 тысяч лет назад? Причем у африканских линий группы А можно косвенным путем, а именно при сравнении протяженных гаплотипов с «неглубокими» общими предками, достичь времен жизни общих предков 132 тысячи лет назад, а у неафриканцев, как их гаплотипы ни комбинировать и ни сравнивать, глубже 64±6 тысяч лет уйти не получается. Более древних общих предков как корова языком слизнула. Что же это за корова была такая?
Некоторые предположения в цитированной выше статье сделаны были – упомянуто возможное последствие извержения вулкана Тоба в Индонезии, около 70 тысяч лет назад. Не исключено было резкое похолодание, возможно, как результат извержения того же вулкана, причем, по мнению специалистов, ранг катастрофичности глобального похолодания намного выше, чем извержения вулкана, но это все оставалось довольно абстрактными предположениями. И вообще – чтобы извержение вулкана в Индонезии привело к гибели почти всех людей на планете – это кажется маловероятным, но все это «по понятиям». Нужно было мнение специалиста, пусть мнение и спорное, неоднозначное, но главное – подтвержденное конкретными данными.
И такой специалист по данной проблеме есть. Знакомьтесь – Валерий Павлович Юрковец, профессиональный геофизик, палеоклиматолог, специалист в области влияния климатических катастроф, в том числе импактного характера, на миграции древнего человека, член Академии ДНК-генеалогии с начала ее образования несколько лет назад.
Продолжение следует…
Анатолий А. Клёсов,
доктор химических наук, профессор
Осталось подойти к решению сформулированной в статье (Klyosov, Rozhanskii, 2012) загадки – повторим ее опять: куда же делись предки Y-хромосомных линий в Африке ранее 5-10 тысяч лет назад, и у неафриканцев ранее 64±6 тысяч лет назад? Причем у африканских линий группы А можно косвенным путем, а именно при сравнении протяженных гаплотипов с «неглубокими» общими предками, достичь времен жизни общих предков 132 тысячи лет назад, а у неафриканцев, как их гаплотипы ни комбинировать и ни сравнивать, глубже 64±6 тысяч лет уйти не получается. Более древних общих предков как корова языком слизнула. Что же это за корова была такая?

Некоторые предположения в цитированной статье сделаны были – упомянуто возможное последствие извержения вулкана Тоба в Индонезии, около 70 тысяч лет назад. Не исключено было резкое похолодание, возможно, как результат извержения того же вулкана, причем, по мнению специалистов, ранг катастрофичности глобального похолодания намного выше, чем извержения вулкана, но это все оставалось довольно абстрактными предположениями. И вообще – чтобы извержение вулкана в Индонезии привело к гибели почти всех людей на планете – это кажется маловероятным, но все это «по понятиям». Нужно было мнение специалиста, пусть мнение и спорное, неоднозначное, но главное – подтвержденное конкретными данными. И такой специалист по данной проблеме есть. Знакомьтесь – Валерий Павлович Юрковец, профессиональный геофизик, палеоклиматолог, специалист в области влияния климатических катастроф, в том числе импактного характера, на миграции древнего человека, член Академии ДНК-генеалогии с начала ее образования несколько лет назад. В.П. Юрковец написал статью в сентябрьский выпуск Вестника Академии ДНК-генеалогии, и я здесь кратко перескажу ее основные результаты и выводы, с моими дополнениями по ДНК-генеалогии.
В.П. Юрковец приводит убедительные доказательства того, что между 57 и 71 тысяч лет назад крупное космическое тело упало в Тихий океан, что, в свою очередь, вызвало мегацунами, так называемое космогенное мегацунами. Размер этого тела был, вероятно, сопоставим с Ладожским импактом – падением на север Русской равнины массивного астероида около 11 километров в поперечнике, но по катастрофическим последствиям превосходящим его на порядки. Как пишет В.П. Юрковец, каменный метеорит диаметром 100 м вызывает на суше разрушения в радиусе немногим более километра, что моментально приводит к возникновению очень сильного цунами сейсмического происхождения. Но при падении крупного космического тела в океан разрушительные последствия неизмеримо более сильные. При этом возникают несколько следующих друг за другом гигантских волн.
Первая – от непосредственного удара о водную поверхность, вторая – от схлопывания (заполнения водой) сформированного ударом водного кратера (каверны), третья – от кратерообразования в океанском дне, то есть в толще пород, слагающих дно океана, и от сопутствующих вторичных процессов. При диаметре метеора 11 км он превосходит по размерам глубину океана практически в любом месте, достигает океанского дна в момент удара, и без труда может пробить тонкую (средняя толщина 7 км) океаническую кору, что приведет к извержению и возникновению дополнительной волны цунами. В открытом океане цунами распространяются со скоростью, достигающей 700 км/час, то есть скорости самолета. Космогенные мегацунами образуют волны, достигающие в высоту нескольких километров, и перехлестывающие самые высокие горные хребты. Это не выдержки из фантастических произведений, это описание того, что на Земле уже происходило, и к тому есть доказательства, в виде многочисленных фотографий последствий этих явлений и соответствующих расчетов, во множестве приведенных в статье В.П. Юрковца. Мегацунами – вот что «как корова языком слизнула» человечество в середине 70-го тысячелетия до нашей эры, 64±6 тысяч лет назад, как показывает ДНК-генеалогия, что совпадает с 4-й морской изотопной стадией, или MIS 4 (71-57 тыс. лет назад), выделяемой палеоклиматологами по резкому ухудшению климатических условий.
Но там было не только резкое ухудшение климатических условий, ухудшение может быть по разным причинам. И извержение вулкана Тоба около 70 тысяч лет назад может случайно (или не случайно) совпасть по времени с исчезновением почти всего человечества. То, что именно в то время на палеоклиматических кривых, приведенных в статье В.П. Юрковца, наблюдается резкое увеличение пыли в атмосфере, понижение температуры, увеличение массы ледников – это, конечно, человечеству не подарок, но о мегацунами пока не говорит. А вот тот факт, что в дополнение ко всему этому палеоклиматические кривые свидетельствуют о резком засолении всей Антарктиды, то есть захлестывании всего материка морской водой, это уже аномалия глобального характера. Тем более что ледовые керны на обоих полушариях показывают резкое увеличения натрия во льду, который мог поступить только с морской водой. Это более чем экстраординарное событие произошло 68 тысяч лет назад. Таким образом, как подчеркивает В.П. Юрковец, все катастрофические события того времени оказались связаны в единый узел.
Еще о катастрофичности тех событий. Судя по размерам кальдеры Тоба (около 100 километров по длинной оси), космический толчок, спровоцировавший его извержение, не уступал по мощности Ладожскому событию, извержение которого сформировало собственную кальдеру оседания около 130 километров в диаметре и спровоцировало извержения вулканов, как минимум на Кавказе и в Италии. Как уже отмечалось, Ладожский астероид имел в поперечнике около 11 километров (Юрковец, 2012), и космическое тело, породившее мегацунами в Тихом океане, вероятно, не уступало ему в размерах. По крайней мере, вулканические последствия двух падений сопоставимы. Диаметр Ладожской астроблемы составляет больше 100 километров. При диаметре астероида, превышающем глубину океана, слой воды не оказывает существенного влияние на формирование кратера, следовательно, размер кратера, образованный падением астероида, вызвавшего мегацунами в Тихом океане, не может быть меньше нескольких десятков километров в поперечнике. Это уже величина, вполне различимая на картах океанического дна Google, и об этом речь пойдет ниже.
Космогенное мегацунами оставило неизгладимые следы на континентах в виде гигантских знаков ряби, называемых у специалистов «гигантская рябь течения», а также «ламинарных супертечений» и «турбулентных сверхпотоков». Я приведу здесь лишь несколько характерных фотографий из статьи В.П. Юрковца, у него их там много десятков. Существует теория дилювиального морфолитогенеза (разработанная доктором географических наук А.Н. Рудым), для гигантских потоков, вызванных прорывами ледниково-подпрудных озёр, и она иллюстрируется на активных русловых формах рельефа высотой от 2 до 20 метров. Это, конечно, ни в какое сравнение не идет с рельефами, вызванными мегацунами 57-71 тыс. лет назад на всех континентах планеты, как показано ниже.
Гигантские знаки ряби течения – это система вытянутых гряд или цепи дюн серповидной формы, синхронно ориентированных навстречу образовавшему их потоку, иногда разделенных перемычками, сопоставимых с высотами основных дюн. Специалистам известны и углы образования проксимальных и дистальных склонов гряд ряби течения и дюн, их профилей, пригребневых частей, и так далее. По ним на аэро- и космоснимках определяют направления дилювиальных потоков прошлого. Типичные размеры гигантских знаков ряби русловых форм рельефа – высота от 2 до 20 м при длине волны (растояние между цепями дюн) от 5-10 м до 300 м. Вот, например, аэрофотоснимок классической гигантской ряби течения в Курайской впадине на Алтае (Рудой, 2006), приведенный В.П. Юрковцом в своей статье:

А вот уже следы космогенного мегацунами, на многочисленных примерах, исследованных В.П. Юрковцом на разных континентах. Это – пустыня Намиб в юго-западной Африке. Те же характерные морфологические признаки, те же гряды, те же перемычки, то же соотношение высоты дюн и максимальной длины волны – только на порядок выше, чем для гигантской ряби течения более традиционных «мегапотоков». В пустыне Намиб высота дюн – до 200 м, длины волн – до 3 километров. Средние величины – 150 м и 2.2 километра, что опять повторяет «классическое соотношение». Здесь и далее – Google maps, за исключением наземных фотографий.

Следы космогенного мегацунами – гигантская рябь течения в пустыне Намиб в юго-западной Африке (из статьи В.П. Юрковца). При увеличении картина выглядит еще более впечатляюще – протяженность дюн может достигать 20 километров:

Не нужно здесь обманываться словом «дюны», это не песчаные дюны, они только присыпаны песком. Основа дюн – грубообломочный (дилювиальный) материал, крупный щебень, только мегаразмера, опять мега, других измерений не подобрать.

Вот так дюны выглядят:

Ориентация дистальных склонов дюн – на восток, то есть поток пришел со стороны моря. Мощные водные потоки выработали много характерных узких каньонов с многочисленными следами кавитации на стенах:

Кавитация – это следы взрывного схлопывания каверн, образующихся при перепадах давления в жидкости, движущейся с переменной скоростью. Вследствие свойства несжимаемости, фронт ударной волны, образующийся при схлопывании каверны, имеет большую крутизну и подобен взрыву. Поэтому кавитация обладает большой разрушительной силой, она оставляет особые следы на материале, на который воздействовал скоростной поток. К ним относятся многочисленные следы «микровзрывов» – поверхности сколов в дилювии и материале, контактирующим с потоком, движущимся с большой скоростью. Ни один другой агент – ветер, попеременно замерзающая и оттаивающая вода, обычное течение, волно-прибойная деятельность и пр. не оставляет подобных следов.
По расчетам, глубина (высота) потока, накрывшего пустыню Намиб, могла составлять от 9 до 15 километров. Эти и другие расчеты показывают, что следы гигантского цунами, которые присутствуют на всех материках, были оставлены несколькими последовательными волнами, образованными единым падением массивного космического тела в Тихий океан:

Проявления космогенного мегацунами многочисленны. Это, например, вортексы («исполинские котлы»), выработанные гигантскими водоворотами, несущими камни, и достигающие десятков и сотен метров в диаметре, хотя в Северной Африке есть и гигантский вортекс диаметром около 40 километров.

Эрозионно-кавитационные образования – вортексы («исполинские котлы»), выработанные гигантскими водоворотами, несущими камни. Вортекс на фото – диаметром 80 метров, в Австралии. Фото В.К.Гусякова (Алексеев, Гусяков, 2011). Вортекс в Мавритании, Северная Африка:

Это также геоскульптурные эрозионно-кавитационные формы рельефа бассейна реки Колорадо

на восточных склонах Кордильер и Скалистых гор

в высокогорных регионах Анд в Южной Америке






на Дальнем Востоке (Китай), в Сибири (Красноярские столбы), на Урале (плато Маньпупунёр)



Но вернемся к волнам гигантской ряби. Итак, следы космогенного мегацунами на всех континентах показывают, что высота гигантской волны, возникшей в результате падения крупного космического тела в Тихий океан, достигала высоты до 15 км. Она преодолевала высочайшие горные хребты и даже континенты. Встречные волны ударились друг о друга в районе северной Африки, именно там зафиксирована хаотичная картина гигантских знаков ряби, «супертечений», «мегапотоков» и прочих свидетельств движения огромных масс воды. Наиболее близкой к центру мегацунами оказалась Австралия, где оставленная в пустыне Симпсона гигантская рябь течения отличается наиболее строгой геометрией, из всех встречавшихся:

Здесь длина волны гигантской ряби составляет около 300 метров. Протяжённость дюн – до 20 и более километров. И эта единообразная картина наблюдается по всей пустыне, протяженностью более 500 километров. То есть это был ровный, без внутренних флуктуаций и завихрений, единый и невероятно грандиозный поток, который, вероятно, даже «не заметил» Австралию – предыдущая тысяча километров суши, включая среднегорье, по которой он прошёл до пустыни Симпсона, никак не сказались на его «ламинарности».
В Северной Америке мегацунами перехлестнуло Кордильеры и Скалистые горы и фотографии разрушений на противоположной от океана, восточной стороне гор, приведены выше. В пустыне Хила, расположенной в районе американо-мексиканской границы на территории американских штатов Аризона и Калифорния и мексиканского штата Сонора, к северу от Калифорнийского залива, оно оставило гряды гигантских дюн – гигантскую рябь течения с высотой дюн до 150 м и длиной волны, превышающей 4 километра – на километр больше, чем в пустыне Намиб. Морфология дюн такая же в обеих пустынях – в Африке и Северной Америке. Пройдя дальше, в штате Колорадо, суперпоток быстро преодолевал турбулентность при выходе из долины реки Литл Колорадо, и создал рельеф, для которого даже нет названия в существующей терминологии ландшафтов – дюны в виде широких ячеистых полос и переходом в протяженные гребни, разделенные впадинами:

В одном месте по ходу движения поток встретил препятствие – одиночную вершину, и вот к чему это привело:

Здесь физико-математическая реконструкция выявила действие трех ударных суперпотоков, причем первый поток перехлестнул через вершину, высота второй волны была меньшей, третьей волны – еще меньшей, она породила турбулентные потоки, отходящие непосредственно от подножия горы. В итоге долины, отходящие от горы, имеют вихревую природу. Еще одно поле дюн турбулентности находится в Аризоне, его размер по диагонали составляет около 4 километров:

В Южной Америке волнам мегацунами противостояли величайшие горные хребты мира, к тому же параллельные друг другу и поперек удару суперпотока. Но это не стало помехой – поток просто перехлестнул их и к тому же пропилил в самых высокогорных частях Анд целую систему долин-спилвеев и изменил направления рек, которые стали течь тоже поперек хребтов (верховья Рио-Гранде и других рек). Примеры геоскульптурных образований приведены выше, на фотографиях в высокогорных регионах Анд в Южной Америке. Там даже нет окатанного галечника, что свидетельствует о кратковременности потока. Гигантские ряби течения встречаются, например, в Аргентине, на плато Альтиплано, и на равнинных территориях, которые даже не закрываются сплошным лесом. Там средняя длина волны превышает 4 километра, что показывает, что удар мегацунами по Южной Америке был сильнее, чем по Северной Америке.

В Азии мегацунами оставило многочисленные следы в виде гигантской ряби – например, в восточной части пустыне Гоби, со средней длиной волны 3.4 километра:

На западной части пустыни Гоби длина волны достигала уже 3.2 километра, то есть поток несколько ослабевал при продвижении на запад. В Монголии, в долине реки Завхан, длина волны уменьшилась уже до полутора километров:

В районе Каспия картина следов мегацунами стала более хаотичной, но вал с огромной скоростью катился дальше, в Европу, которая, видимо, пострадала меньше всех. Южный рукав мегапотока ударил по Памиру и Гималаям, образовав классическую гигантскую рябь течения в пустыне Такла-Макан, с длиной волны до 3 километров:

При этом отраженные волны мегацунами встретились, что привело к образованию серии «дюн турбулентности», направленную дистальными склонами на северо-восток:

По ходу своего движения поток пропилил, наверное, самый большой в мире спилвей – Алайскую долину, отделившую Памир от Гиссарского и Зеравшанского и Алайского хребтов, по которой вода устремилась в гигантский бассейн Каспийского и Аральского морей, перехлестнул через горы на Иранское плато, и сформировал там сразу несколько спилвеев. Пример – ущелье Танге Ваши, неподалеку от современного Тегерана:

Следы мегапотока в Индии-Пакистане, в пустыне Тар, протяженностью 850 км (северо-запад Индии и юго-восток Пакистана), где зоны турбулентности чередуются с ламинарным течением:

Гигантская рябь течения встречается на юго-востоке Аравийского полуострова, откуда мегаволна ударила по Африке, пройдя континент с юга на север. Пример ламинарного супертечения к западу от низовий Нила:

А вот – и более обширная картина в Африке:

Еще доказательство мегацунами – солёные бессточные высокогорные озера. Самое высокое в мире – соленое бессточное озеро Намцо в Тибете, высота над уровнем моря 4718 метров. В попытках объяснить это явление была выдвинута гипотеза, что соль поступала с дождевыми водами, текущими по горным породам, хотя воды текут везде, но озера почему-то пресные. Бессточное озеро Балхаш (высота над уровнем моря 340 м), например, состоит из двух частей – соленая восточная, и пресная западная, соединенные небольшим проливом. В пресную впадает река, которая несет соли, но это не дает особой солености. Отчего другая часть почти на порядок более соленая – никто не объясняет, и в Википедии кроме описания этого факта, а также что соленость пресной части 0.74 г/л, а соленой части 3.5-6.0 г/л, больше ничего нет. В.П. Юрковец относит солёность Балхаша за счет реликта некоего события, которое с поступлением соли из окрестностей никак не связано. Какое это могло быть событие – примерно уже ясно, хотя бы как вариант.
Наконец, на последствия космогенного мегацунами могут указывать океанские морфологические структуры, образованные, видимо, мощнейшими потоками, которые буквально за минуты и часы вырабатывали глубоководные долины, каменный материал которых тут же сбрасывался в океанический простор. Пример – южный прорыв между Южной Америкой и Антарктидой

Северный прорыв, состоящий из трех шлейфов, возможно, трех волн космогенного мегацунами, между Южной и Северной Америками:

При этом сформировались глубоководные желоба (на снимках выделяются темно-синим, почти черным цветом), находящиеся на фронтальной части дуг (шлейфов), сформированных обломочным материалом, предельные глубины которых составляют между Южной Америкой и Антарктидой 8428 метров, между Америками 8385 метров. Глубже их только впадины, образованные столкновением тектонических плит. Эти желоба были сформированы (вынесены) классическим делювием (тем самым обломочным материалом), который принесен потоком, но не окатан, как показано на фотографии выше.
Полагаю, приведеные здесь данные (они, повторяю, все взяты из статьи В.П. Юрковца в Вестнике Академии ДНК-генеалогии) убедительно свидетельствуют о масштабе космогенной катастрофы, происшедшей на Земле 57-71 тыс. лет назад. Она не могла оставить на планете ничего живого, но, получается, что оставила. Сохранились животные и птицы, хотя известно, что после 50 тысяч лет назад (грубая оценка) большинство животных исчезли с лица Земли. Объяснений нет, разве что «по понятиям», например, что их уничтожил человек. Но сам человек после космогенной катастрофы сохранился в столь небольших количествах, что выживание его висело буквально на волоске.
Рассмотрим сначала неафриканцев. Потомство выжило у минимального их количества, что и привело к эффекту «бутылочного горлышка популяции» 64±6 тысяч лет назад. Фактически, в те времена у неких отца с матерью выжили как минимум двое сыновей, которые тоже дали выжившее потомство, и оно постепенно пошло в количественный рост. Все остальные выжившие после катастрофы люди тоже могли дать потомство, но оно пресеклось, не выжило. В итоге практически все мужчины современности на планете, во всяком случае, те сотни тысяч человек, которые прошли тест на гаплотипы-гаплогруппы Y-хромосомы, сходятся по мутациям в гаплотипах формально к одному общему предку, тому самому патриарху, от которого выжили как минимум двое сыновей, давших выжившее же мужское потомство в середине 70-го тысячелетия назад.
Следует подчеркнуть, что тот патриарх имел в своей Y-хромосоме, разумеется, всю цепочку из сотен тысяч снипов, ведущую от общих предков с приматами, и далее вверх по цепочке снипов Y-хромосомы, через A0-T, A1, A1b, ВТ, как описано в первой части. Носители этих снипов как терминальных погибли в катастрофе, но выживший, ставший патриархом неафриканского человечества, их сохранил. Согласно расчетам компании YFull (http://www.yfull.com/tree/), гаплогруппа ВТ образовалась примерно 126 тысяч лет назад (доверительный интервал 121-132 тыс. лет назад), то есть задолго до катастрофы. Возможно, расчеты потом уточнятся, но примем их за основу, других всё равно нет. Так вот, выживший патриарх имел именно эту гаплогруппу, ВТ. Наверняка сотни тысяч и миллионы погибших в катастрофе имели многие другие гаплогруппы и субклады, образовавшиеся за 60 тысяч лет от образования BT до времени катастрофы, но они погибли, и те гаплогруппы и субклады не были унаследованы. Поэтому отсчет выживших поколений пошел именно от гаплогруппы ВТ, и не от времени ее образования, а от времени бутылочного горлышка, вызванного катастрофой. Эта гаплогруппа и была названа бета-гаплогруппой, по факту выживания, а не по факту образования за 60 тысяч лет до катастрофы.
В отношении женщин пока неизвестно, возможно, их после катастрофы по каким-то причинам осталось намного больше, только толку в том для будущих поколений было немного, если мужчины почти все погибли. Но не исключено, что мужчины с той же Y-хромосомой, то есть от одного отца, родились от многих разных матерей, как и их дочери, которые Y-хромосому не имеют. В таком случае общий предок мтДНК, то есть праматерь, может оказаться значительно древнее, чем 64±6 тысяч лет назад. Но подобные расчеты мне неизвестны, да и методы расчетов по мтДНК не столь совершенны, и продолжают дебатироваться. Тем не менее, в недавней статье (Groucutt et al., Evolutionary Anhropology, 2015) сообщается, что все мтДНК современных неафриканцев сходятся к общему предку, жившему (точнее, жившей) в интервале 60-79 тысяч лет назад. По другим источникам – 60-70 тысяч лет назад (ссылка).
Это – гаплогруппа L3. На схеме ниже (Behar et al., 2012) видно, что старейшая гаплогруппа L0, которую считают «африканской», и которая представляет большую серию из полусотни «африканских» гаплогрупп, найденных в основном к югу от Сахары – среди койсанского населения Южной Африки, а также в Эфиопии, Танзании, Мозамбике и среди пигмеев – не является предковой по отношению ко всем остальным гаплогруппам мтДНК. Более того, время жизни общего предка мтДНК практически совпадает со временем жизни Y-гаплогруппы BT и с временем описываемой здесь космогенной катастрофы.

Наконец, ископаемые неандертальцы в Азии и на Ближнем Востоке тоже датируются не ранее 50-70 тысяч лет (Groucutt et al., 2015), причем сообщается, что все те, для кого были тестированы ДНК, имеют общего предка в том же временном интервале. Иначе говоря, их «разнообразие» было минимальным, все – достаточно близкие родственники.
Возвращаемся к мужской гаплогруппе В. Непонятно, почему в списке YFull время образования гаплогруппы B указано как 83900 лет назад, как и время образования «параллельной» СТ (с доверительными интервалами у обеих от 79700 до 88100 лет назад). С одной стороны, они могли образоваться еще до катастрофы, от той, ранней гаплогруппы ВТ, и уйти, скажем, в Африку, где и выжили, и дали свое потомство. Но гаплогруппа ВТ у «автохтонных африканцев» не обнаружена, да и СТ к Африке никак не относится, тем более с нисходящими гаплогруппами С и F (обе по данным YFull образовались примерно 65 тысяч лет назад, с доверительным интервалом между 62 и 68 тысяч лет), и у каждой много нисходящих снипов, явно не африканских. Да и дерево гаплотипов человечества, приведенное в первой части этой статьи, показывает, что гаплогруппа В образует единую систему с неафриканскими гаплогруппами.
Вывод – что-то с датировкам В и СТ у YFull не то. Они не могли быть ранее 64±6 тысяч лет назад. Правда, взгляд на количество снипов, по которым предположительно YFull должны были вести расчеты, объясняет, в чем у YFull проблемы. Для одинаковой датировки гаплогрупп В и СТ у них должно быть одинаковое число снипов, а там стоит 30 снипов у гаплогруппы В, и 316 снипов у гаплогруппы СТ. Иначе говоря, по данным компании YFull (144 лет на снип), гаплогруппа В должна отстоять от ВТ на 4300 лет, а СТ – на 45 тысяч лет. Это с расчетами YFull с никакой стороны не сходится, в том числе и с постулатом равного количества лет для «параллельных» снипов. Поэтому вопрос пока закроем до пересчетов компанией YFull своих (точнее, чужих) данных.
Итак, с гаплогруппами, и вообще с ДНК-генеалогией человечества, в целом ясно, как и с возрождением его, человечества, после космогенной катастрофы 64±6 тысяч лет назад. Это, правда, ставит другую потрясающую загадку – что же представляло человечество до катастрофы? Какие там были цивилизации? Что они после себя оставили, что не было смыто мегацунами? Понятно, что самих людей не осталось, они были разорваны в клочья ударами волн в смеси с острым щебнем, высотой до 15 км и несущимися со скоростью самолета, перехлестывающих через самые высокие горы мира. Остались только немногочисленные скелетные остатки, с которыми работают палеоантропологи, да и те древние люди в большинстве своем определенно не являются нашими предками, чисто статистически они были предками погибших в космогенной катастрофе. Поэтому сопоставления «архаичности» ископаемых скелетных остатков с антропологией современного человека, скорее всего, являются безотносительными. Это покажет тестирование этих «архаичных» на ДНК, откуда потоком пойдут неизвестные нам гаплотипы и гаплогруппы, во всяком случае после A0-T, А1 и A1b. Но может, и тех не будет.
Излагаемая здесь новая концепция затрагивает и другие загадки, давно волнующие исследователей. Например, загадкой является, как получилось, что предки современных людей совершили столь быстрый переход от каменного века до более современных технологий. Сейчас становится понятным, что люди, выжившие после катастрофы, и выброшенные природой обратно в каменный век, были людьми ментально относительно развитыми, после ста тысяч лет цивилизации, нам, впрочем, пока неизвестной. И свои природные способности они относительно быстро возвращали в ходе тысячелетий выживания после катастрофы, следуя той хрестоматийной экспоненте прогресса человечества.
Еще одна загадка, точнее, комплекс загадок – это находимые время от времени древние объекты и технологии, которые не находят объяснения в рамках существующих представлений, и которые, более того, не могут быть воспроизведены. Это и способы перемещения гигантских каменных блоков, и изготовления блоков пирамид с помощью предполагаемых технологий древности, как, впрочем, и укладка самых древних пирамид, встык, без зазоров на гладкой поверхности, из огромных многотонных блоков, и древнейшие пирамиды из странных материалов, нам сейчас неизвестных, и многое другое. Обычно люди, которые пытаются обсуждать это вопросы, подвергаются осмеянию людишками, к решению сложных вопросов вообще неспособными. Но так устроен современный мир, в котором подавляющее большинство критиков и отвергающих новые выдвигаемые концепции, причем отвергающие «по понятиям», являются мизерными посредственностями. «Фрик» – любимое их словечко.
Перейдем теперь к африканской части человечества. Впрочем, всё, что сказано выше о цивилизациях во времена до космогенной катастрофы, относится и к африканцам, если они там жили до 60 тысяч лет назад. Мы ведь и этого не знаем. Времена общих предков современных «автохтонных» африканцев не превышают 5-10 тысяч лет назад (см. первую часть), а цепочки их снипов, уходящие на сотни тысяч лет назад, не привязаны к Африке. Тем не менее, будем исходить из того, что люди в Африке жили в период до катастрофы, ведь находят же там древние скелетные останки людей с датировками до 200 тысяч лет назад. Мы, повторяю, не знаем, были ли они нашими предками, и не узнаем, пока их кости не будут тестированы на ДНК. Так вот, подавляющее большинство тех людей тоже погибли в космогенной катастрофе, но в отличие от неафриканцев выжили люди, возможно, всего несколько человек, но из разных ДНК-линий. Неафриканец-патриарх, напоминаю, был гаплогруппы ВТ. Патриархи-африканцы были нескольких линий, но мы не знаем, какие они были, где те линии/гаплогруппы образовались, возможно, вне Африки, и их носители пришли туда позже, но до катастрофы. А может, и после катастрофы.
Поскольку крупное космическое тело упало в Тихий океан, как показано на карте-схеме выше, и самым сильным волновым ударам подверглись обе Америки, Азия и Австралия, то Африка оказалось несколько в стороне и на большем расстоянии. Разрушения и там были гигантскими, но какое-то число носителей этих пока неизвестных нам гаплогрупп выжили, вместе с женщинами, носителями наследственных мтДНК. Поэтому мужчины начали свое возрождение не с одной гаплогруппы, а с нескольких. Там самым они получили некоторое «разнообразие», которое впоследствии многократно приумножилось за счет многочисленных миграций в Африку – особенно прибывших гаплогрупп В, E1b, R1b (в Камеруне-Чаде и Южной Африке, потомков буров), J1 и J2 (включая племя Лемба в Южной Африке и им подобных), и так далее. Так в Африке сформировалось «генетическое разнообразие», которое к «автохтонным» африканцам практически не относится. Африка – открытая система, а понятие «разнообразие» к открытым системам не относится.
Хочется верить, что в черной Африке тоже были продвинутые древние цивилизации, но пока свидетельств мало, кроме тех, что на севере Африки, в частности, древнейшие пирамиды и Сфинкс. На них ясно видны кавитационные разрушения и знаки водной эрозии на большой высоте, которые традиционно приписывают действию ветра и песка. В.П. Юрковец придерживается другой точки зрения, и не без оснований. Во всяком случае, его точка зрения должна быть рассмотрена на равных, по гамбургскому счету. Однако про критиков было выше.
Анатолий А. Клёсов, доктор химических наук, профессор
Но разбираться особенно и не надо, потому что у всех мужчин на Земле, африканцев и неафриканцев, во всяком случае, у всех, кто делал тест на ДНК, снип-цепочки Y-хромосомы по древности одинаковы. У всех эти цепочки снипов тянутся от общих предков с шимпанзе, орангутангом, гориллой, макакой и другими древними общими с нами предками. Ниже – характерный пример, процитированный еще в 2012 году в журнале Advances in Anthropology (Klyosov, Rozhanskii, Ryabchenko):
человека, шимпанзе, гориллы, орангутанга и макаки
Видно, что в 90% случаев нуклеотидные последовательности показанных фагментов Y-хромосомы совпадают. А у шимпанзе и человека совпадают 93 нуклеотида из 97, то есть в 96%. Так что у всех нас ДНК-корни древние, и африканцы в этом отношении никак не выделяются.
Хорошо, а насколько древние? Антропологи оценивают время расхождения линии будущего человека от линии будущего шимпанзе в 4-6 миллионов лет назад, это же повторяют и генетики, хотя поиски статей с конкретными расчетами результатов не дали. А вот результаты расчета с помощью автоматического калькулятора Килина-Клёсова (ККК, 2015), иллюстрация увеличивается по клику:
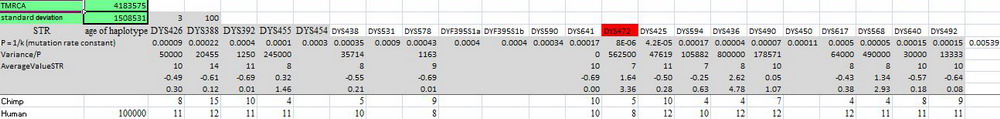
Общий предок человека и шимпанзе жил, если округлить расчеты, 4.2±1.5 миллионов лет назад. 16-маркерный гаплотип шимпанзе был извлечен из геномной базы данных, как описано в той статье в Advances in Anthropology (2012), а в качестве обобщенного гаплотипа человека брался экстраполированный предковый гаплотип по всем неафриканским гаплогруппам, пересчитанный на времена 100 тысяч лет назад. Впрочем, можно было и не пересчитывать, вот – время до общего предка современных носителей гаплогруппы А00 и шимпанзе, иллюстрация увеличивается по клику:

Оно равно 4.4±1.4 миллионов лет назад, что практически одинаково в пределах погрешности расчетов. Те же времена дают все основные гаплогруппы серии А, например:
• А0а-V150 имеет общего предка с шимпанзе 4.6±1.7 млн лет назад;
• A1b1-M32 (Чад и Турция) 4.3±1.4 млн лет назад;
• A1b1-M32, M13 (Британия) 4.2±1.4 млн лет назад, а также
• гаплогруппа В, 4.4±1.4 млн лет назад.
Короче, в отношении «древних ДНК-линий» африканцы ничем от неафриканцев не отличаются. И те, и другие тянут свои ДНК-линии от общего предка с шимпанзе. И у тех, и у других цепочки снипов уходят на глубины в сотни тысяч и миллионы лет. К этому мы еще вернемся.
Может, под «древними африканскими ДНК-линиями» попгенетики и прочие генетики подразумевают древние времена жизни их общих предков, и полагают, что они намного более древние, чем у неафриканцев? Придется их огорчить, ситуация противоположная. Африканские предки группы А (популяций, субкладов) почти все совсем недавние. Возьмем, например, гаплогруппу А00, которая среди гаплогрупп считается самой древней. Да, для нее цепочка снипов насчитывает – по данным, приводимым YFull – 1684 снипов, что при 144 годах на снип (это число приводит YFull) дает 242500 лет от начала гаплогруппы. Действительно, YFull приводят близкий «возраст» А00, 235 тысяч лет. Но откуда эта цепочка снипов ведет – никто не знает. Может, из Африки, может, с Кавказа, может, из Сибири, может, из Европы – неизвестно. Почему-то считается, что именно из Африки. Но это опять «по понятиям», правда, откуда появившимся, непонятно, извините за игру слов. Древнейшие ДНК находили в Европе, на Русской равнине, в Сибири, а в Африке ни разу, правда, и исследований таких не было. Так откуда эти «понятия» появились? А так, стиль попгенетиков.
Кстати, о возрасте гаплогруппы А00. Калькулятор КК дает для базовых гаплотипов А0 и А00 возраст общего предка (с округлением) 217000±87000 лет. Это – возраст А00 или немного старше. Так что датировка по снипам воспроизводится по гаплотипам в пределах погрешности расчетов (иллюстрация ниже увеличивается по клику).

Так вот, об общих предках африканских популяций, субкладов группы А. Они почти все по времени очень мелкие, часто всего несколько сотен лет. Иногда – между 1000 и 2000 лет. Самые древние – примерно 5000-7000 лет. И опять – где эти предки жили, никто не знает. Взглянем на гаплогруппу А00. В литературе (Mendez et al., 2013) описаны 11 гаплотипов в 95-маркерном формате (без 16 мультимаркерных гаплотипов) носителей гаплогруппы А00, все из племени Mbo из Камеруна, и еще один – афроамериканец. Расчетов в статье, конечно, нет. Все 11 гаплотипов содержат 29 мутаций, что дает 29/11/0.161 = 16 условных поколений, но с учетом того, что 45 аллелей (из 1045 аллелей) авторы определить не смогли, то до общего предка на самом деле 17 условных поколений, то есть 425±90 лет.
Итак, сама гаплогруппа А00 образовалась более 200 тысяч лет назад, причем неизвестно где, а общий предок носителей А00 из Камеруна жил примерно 425 лет назад. Если добавить к серии из Камеруна еще гаплотип афроамериканца той же гаплогруппы, который отличается от предкового гаплотипа камерунцев на 11 мутаций, то есть на 11/0.161 = 68 → 73 условных поколения, то есть на 1825 лет, то их общий предок, камерунцев и афроамериканца, жил примерно (425+1825)/2 = 1125 лет назад.
Примеров недавних общих предков разных мужских популяций гаплогрупп класса А, которые сейчас живут в Африке, можно привести немало. В большинстве этих популяций гаплотипы или одинаковы или почти одинаковы, даже для протяженных гаплотипов. С доступными гаплотипами гаплогруппы А00 мы уже разобрались, им четыре сотни лет. Взглянем теперь на Проект FTDNA гаплогруппы А:
• В гаплогруппе/субкладе А0-V150 имеются два 25-маркерных гаплотипа, оба идентичны друг другу, то есть их общий предок жил не более 300-400 лет назад. Других гаплотипов в 25-маркерном формате или выше нет.
• В гаплогруппе A0a-L981 (L994, V148, V166) между двумя доступными 67-маркерными гаплотипами есть 8 мутаций, что помещает их общего предка на 900 лет назад.
• В гаплогруппе A0b-V166, L896 шесть 37-маркерных гаплотипов имеют общего предка 475±160 лет назад.
• В гаплогруппе A1a-M31 («европейский кластер») пять 37-маркерных гаплотипов имеют общего предка 1175±290 лет назад.
• В гаплогруппе A1a-M31 (все неизвестного происхождения) пять 37-маркерных гаплотипов имеют общего предка 1880±390 лет назад.
• В гаплогруппе A1a-M31 (из 18 гаплотипов – 14 неизвестного происхождения, два из Англии, один из Швейцарии, один Кейп Верде) все 37-маркерные, все имеют общего предка 730±130 лет назад.
• В гаплогруппе A1a-M31 («британский кластер») 19 гаплотипов в 67-маркерном формате имеют общего предка 740±120 лет назад.
В итоге, 50 гаплотипов в 37-маркерном формате четырех предшествующих серий (А0а-М31) имеют общего предка 9270±970 лет назад. Но надо обратить внимание, что это четыре серии гаплотипов, разбросанные по разным континентам. Это – не одна «ДНК-линия», это – четыре разных ДНК-линии, с четырьмя общими предками, каждый «глубиной» от 700 до 1900 лет.
Ну и где, позволительно спросить, «древнейшие ДНК-линии в Африке»? Это – миф, ничем не обоснованный. Ископаемых ДНК нет, древних линий, как мы видим, нет, к тому же, в Африке вовсе не гаплотипы группы А явяются доминирующими. Доминирует гаплогруппа E1b; в Камеруне и Чаде, и отчасти в Нигерии, доминирует гаплогруппа R1b, поэтому неудивительно, что в Африке наблюдается «высокое генетическое разнообразие», только это не от древности, а от обилия носителей разных гаплогрупп, прибывших в Африку в разные времена. Да и вообще, как можно серьезно обращать внимание на «генетическое разнобразие» в открытых системах, какой является Африка? Как я уже не раз отмечал, самым «генетически разнообразным» городом мира является, наверное, Нью-Йорк, но вряд ли кто будет серьезно утверждать, что Нью-Йорк – древнейший город мира. Кроме, наверное, популяционных генетиков, с их примитивными и бездумными критериями.
Откуда же появился этот миф о «древности африканских линий»? Датировок ведь их до последнего времени не было, да и датировки бывают, как мы убедились, нескольких видов – цепочек мутаций, уходящих далеко вглубь, до приматов; цепочек мутаций, ведущих ко временам образования соответствующих гаплогрупп; и датировок общих предков современных носителей соответствующих гаплогрупп и субкладов. Ни одна из этих датировок не «укоренена» территориально, то есть мы не знаем, на какой территории, на каком континенте образовалась та или иная гаплогруппа. Мы можем только набирать сведения и складывать фрагменты общей картины, но Африка в этом отношении находится на одном из последних мест по накопленным знаниям. Но фантазий, натяжек и откровенных фальсификаций про них со стороны попгенетиков – хоть отбавляй.
Собственно, это характерно для популяционных генетиков, но они уроки из принципиальных ошибок не извлекают. Еще не так давно они утверждали, что гаплогруппа R1b образовалась в Европе 30 тысяч лет назад, и сначала приняли в штыки, а потом напрочь игнорировали сведения и датировки ДНК-генеалогии, что R1b прибыла из Южной Сибири длинным миграционным путем в Европу примерно 4800 лет назад (Klyosov, 2009; Klyosov, 2012). Называли это «псевдонаукой», особенно в этом усердствовали попгенетики Балановские. Разворот произошел в 2013 году, когда неподалеку от Байкала нашли ископаемую гаплогруппу R с археологической датировкой 24 тысячи лет назад. Лед тронулся, и попгенетики, не моргнув глазом, не повинившись и не цитируя статьи с датировками ДНК-генеалогии, стали повторять то, что уже было опубликовано нами несколько лет назад. Например, в недавней статье Batini, Jobling и других (2015) датировки гаплогруппы R1b в Европе уже даются как R1b-L11 4510 лет (доверительный интервал 3920-5160 лет назад) и R1b-P312 4210 лет (доверительный интервал 3700-4790 лет назад). YFull дают датировки R1b-U106 4900 лет (доверительный интервал 4400-5400 лет назад), R1b-P312 4600 лет (для TMRCA, доверительный интервал 4300-5000 лет назад).
К сожалению, приходится повторять, что попгенетики уроков из своих ошибок не извлекают. Ситуация повторяется с Африкой. Данных нет, или данные говорят о другом, но их придумывают и искажают. Зачем это им нужно? Для нормального человека звучит невероятно, но это ментальность вассалов, которые лебезят перед сюзереном. Сюзерен решает, кого пригреть, кого пригласить на конференцию, кому дать грант, кого взять в коллективную статью, ввести в комиссию или комитет, в общем, в инфраструктуру. Без сюзерена вассал – никто, и вассал это прекрасно понимает. А зачем это нужно сюзерену? О, там большая политика. Вассала к этим вопросам не допускают. Вассал знает, что если он публично усомнится в африканском происхождении человечества, то его дни сочтены. Поэтому он, вассал, готов разорвать на части любого, кто сомневается или выставляет данные против. А поскольку он разорвать на части не может, то цель – найти компромат, состряпать ложь, передернуть, исказить – других способов или аргументов у вассала нет, во всяком случае, научных.
Посмотрим на схему гаплогрупп и субкладов группы А, приведенную согласно классификации ISOGG, в сокращенном виде:
В этой схеме – источник принципиальной путаницы попенетиков. Они не смогли увидеть и осознать, что эта лесенка субкладов представляет собой серию вилок, которые периодически расходятся на «африканские» (по сегодняшнему месту жительства) и неафриканские ветви. Опишем кратко эти вилки, а потом рассмотрим их подробнее – когда они образовались, и кто являются потомками субкладов по обе стороны тех вилок.
Итак, вилок всего четыре. Первой от ствола, уходящего вглубь эволюционного дерева человечества (точнее, его Y-хромосомы), отходит вилка, знаменующая, видимо, начало Homo sapiens – расхождение на А00 и А0-Т. Вторая – расхождение А0-Т на А0 и А1. Третья – расхождение А1 на А1a и A1b. Четвертая – расхождение A1b на A1b1 и ВТ.
Если упрощенно – то каждая вилка расходится на африканскую (по нынешнему месту жительства) и неафриканскую стороны. А00, А0, A1a и A1b1 уходят в Африку, противоположные стороны вилки A0-T, A1, A1b и BT остаются вне Африки. «Упрощенно» – потому что вилка на А00 и A0-T образовалась еще до расхождения на африканцев и неафриканцев (это расхождение произошло примерно 160 тысяч лет назад), а впоследствии многие неафриканцы перебирались в Африку – это и многочисленные носители гаплогрупп E1b, J2, R1b, и так далее. Среди них – миллионы современных жителей Западной Африки (берберы, жители Камеруна и Чада), египтяне, потомки буров в Южной Африке, и так далее. Немало жителей Европы и Ближнего Востока имеют гаплогруппу A1a, как недавно показывал И.Л. Рожанский в статье на Переформате. Да и вообще носителей группы А в Африке относительно мало – часто на уровне единиц процента или того меньше. Рассмотрим вилки подробнее.
Первая вилка – расхождение предшественника Homo sapiens (или Homo sapiens в начальной стадии, примерно 230 тысяч лет назад) на гаплогруппы А00 и А0-Т. Нисходящих субкладов от А00 пока не обнаружено, и ее нынешние носители обнаружены только в Африке в количестве 11 человек (см. выше), с общим предком несколько сотен лет назад, а также несколько афроамериканцев с одинаковыми гаплотипами. Снипы гаплогруппы А0-Т, напротив, имеются у всех неафриканцев, в том числе у всех нас, читателей Переформата, хотя только у мужчин. И, разумеется, у всех африканцев, носителей гаплогрупп E1b, J2, R1b, В и других, которые прибыли в Африку относительно недавно. Но помимо них снипы группы А0-Т должны быть и у всех африканцев, носителей групп А, потому эти снипы предшествуют разделению африканских и неафриканских линий (примерно 160 тысяч лет назад). Впрочем, речь идет о промежуточных снипах в цепочках наших и африканских снипов, носителей А0-Т как терминальных снипов пока ни у кого не выявлено, ни в Африке, ни вне Африки. Поэтому называть его «африканским» никакого смысла не имеет.
К тому, что у всех нас, мужчин, есть множество снипов гаплогруппы А0-Т – ниже приведен список части из 587 снипов этой гаплогруппы, из которых у меня есть L1090, L1121, L1124, L1125, L1127, L1235 и многие другие:
CTS10042, CTS10627, CTS10834, CTS10847, CTS10976, CTS10993, CTS11148, CTS11184, CTS11411, CTS11468, CTS11530, CTS1169, CTS11720, CTS11734, CTS11853, CTS11913, CTS11948, CTS12010, CTS1211, CTS1277, CTS1278, CTS12947, CTS12972, CTS1340, CTS1367, CTS1415, CTS1416, CTS1417, CTS1419, CTS1422, CTS1619, CTS1738, CTS1795, CTS1907, CTS2134, CTS262, CTS2626, CTS2636, CTS2637, CTS2638, CTS2639, CTS2663, CTS 2680, CTS2891, CTS2988, CTS2992, CTS3067, CTS3075, CTS3229, CTS3230, CTS3231, CTS3316, CTS3321, CTS3358, CTS3402, CTS3441, CTS3527, CTS3534, CTS3548, CTS3551, CTS358, CTS3607, CTS3622, CTS3649, CTS3650, CTS3654, CTS3697, CTS3775, CTS3848, CTS3943, CTS3984, CTS4259, CTS4465, CTS4862, CTS501, CTS5287, CTS5423, CTS5454, CTS5508, CTS5577, CTS5648, CTS570, CTS5936, CTS5979, CTS623, CTS6918, CTS6985, CTS7191, CTS7275, CTS7278, CTS7400, CTS7500, CTS7650, CTS7659, CTS7941, CTS8008, CTS8127, CTS8350, CTS8507, CTS8508, CTS8591, CTS8626, CTS8665, CTS8797, CTS8816, CTS8896, CTS9018, CTS903, CTS9108, CTS9510, CTS9515, CTS9539, CTS9596, CTS9733, CTS9739, CTS9754, CTS9779, F1050, F1209, F1224, F132, F1586, F184, F185, F2957, F3032, F313, F3159, F3166, F3185, F3194, F3197, F3337, F3561, F3564, F3644, F3650, F3692, F4099, F686, F719, F773, F82, F937, F989, IMS-JST002612, IMS-ST022448_3, IMS-ST022448_6, IMS-ST022449_3, IMS-ST022449_6, IMS-ST022451_2, K511, L1002, L1004, L1005, L1009, L1013, L1028, L104, L105, L1053, L1060, L1061, L1062, L1067, L1071, L108, L1089, L1090, L1095, L1098, L1099, L110, L1101, L1105, L1112, L1116, L1118, L112, L1120, L1121, L1123, L1124, L1125, L1127, L1129, L113, L1132, L1135, L1136, L1137, L114, L1142, L1143, L1145, L1150, L1155, L1179, L1185, L1120, L1225, L1235, L132, L1346, L1347, L1352, L1480, L1492, L15, L16, L168, L248, L268, L313, L350, L352, L413, L418, L438, L440, L449, L457, L468, L469, L498, L543, L58, L604, L721, L74, L741, L747, L76, L760, L768, L777, L779, L781, L819, L82, L821, L851, L875, L882, L962, L969, L970, L977, L986, L989, M168, M173, M198, M207, M213, M235, M251, M294, M299, M306, M417, M42, M420, M448, M449, M45, M459, M511, M512, M513, M514, M515, M516, M526, M74, M89, M9, M94, P108, P128, P131, P132, P133, P134, P135, P136, P138, P139, P14, P140, P141, P143, P145, P146, P148, P149, P151, P157, P158, P159, P160, P161, P163, P166, P187, P19_1, P207, P224, P225, P226, P228, P229, P230, P231, P232, P233, P234, P235, P236, P237, P238, P239, P242, P243, P244, P245, P27.1_2, P280, P281, P282, P283, P284, P285, P286, P294, P305, P316, PAGES00026, PAGES00081, PAGES00083, PF1345, PF1378, PF1389, PF1396, PF1402, PF1403, PF1405, PF1407, PF1408, PF1411, PF1412, PF1414, PF1415, PF1418, PF1419, PF2589, PF2590, PF2591, PF2608, PF2610, PF2611, PF2614, PF2615, PF2616, PF2619, PF2620, PF2621, PF2622, PF2626, PF2629, PF2631, PF2634, PF2635, PF2637, PF2640, PF2642, PF2647, PF2651, PF2653, PF2658, PF2660, PF2683, PF2685, PF288, PF2690, PF2694, PF2695, PF2700, PF2709, PF2716, PF2718, PF2722, PF2723, PF2729, PF2731, PF2732, PF2736, PF2737, PF2739, PF2742, PF2747, PF2748, PF2749, PF2752, PF2753, PF2754, PF2755, PF2756, PF2757, PF2758, PF2770, PF2772, PF2988, PF349, PF3494, PF3495, PF3496, PF3497, PF3500, PF4522, PF5459, PF5469, PF5477, PF5483, PF5488, PF5490, PF5498, PF5500, PF5503, PF5513, PF5861, PF5862, PF5863, PF5864, PF5869, PF5870, PF5871, PF5872, PF5876, PF5881, PF5882, PF5885, PF5886, PF5888, PF5889, PF5895, PF5908, PF5911, PF5914, PF5916, PF5919, PF5920, PF5936, PF5940, PF5943, PF5944, PF5945, PF5949, PF5951, PF5953, PF5954, PF5955, PF5956, PF5957, PF5958, PF5964, PF5965, PF5967, PF5970, PF5971, PF5975, PF5978, PF5980, PF5981, PF5982, PF5984, PF5985, PF5991, PF6013, PF6016, PF6047, PF6054, PF6056, PF6057, PF6066, PF6069, PF6071, PF6072, PF6073, PF6114, PF6116, PF6118, PF6121, PF6132, PF6133, PF6143, PF6153, PF6156, PF6158, PF6159, PF6160, PF6162, PF6163, PF6165, PF6168, PF6169, PF6170, PF6172, PF6179, PF6181, PF6194, PF6195, PF6201, PF6204, PF6211, PF6212, PF6213, PF6214, PF6215, PF6216, PF6217, PF6218, PF6220, PF6222, PF6230, PF6231, PF6233, PF6512, PF682, PF7514, PF7522, PF7527, PF7530, PF7531, PF7532, PF7534, PF7540, PF7542, PK1, PR1220, PR202, PR2119, PR5761, PR855, rs161492_1, V221, V238, V241, V250, Y2902, Y32, Y33, Y34, Y36, Y783.2, YSC0000057, YSC0000077, YSC0000081, YSC0000107, YSC0000108, YSC0000109, YSC0000110, YSC0000193, YSC0000350, YSC0000913, Z1331, Z280, Z282, Z5997, Z6030, Z6032.
Понятие «вилка» не стоит понимать буквально, А00 и А0-Т могли образоваться с разницей в десятки тысяч лет. По данным YFull, до начала А00 насчитывается 1684 снипа, до начала А0-Т – 587 снипов, но возраст обеих гаплогрупп почему-то выставлен как одинаковый, 235 тысяч лет назад. На такую повторяющуюся «дихотомию» в презентации данных YFull мы уже не раз обращали внимание. В любом случае, искать у нас, читателей Переформата, снипы гаплогруппы А00 бесполезно, нет их у нас, ни одного из тех 1684 снипов. Но важно то, что А0-Т образует очередную вилку, на гаплогруппы А0 и А1.
Вторая вилка – расхождение А0-Т на А0 и А1. Потомки А0 сейчас живут в Африке, потомки А1 – мы с вами, читатели Переформата. У всех нас в цепочках снипов есть снипы субклада А1 (см. ниже), но это опять не значит, что наши предки вышли из Африки. Это опять промежуточные снипы, и никого с терминальными снипами А1 пока не выявлено, ни в Африке, ни вне Африки. Поэтому называть его «африканским» никакого смысла тоже не имеет.
По данным, приведенным YFull, к A0 ведет 899 снипов (это 129460 лет, если снип образуется в среднем раз в 144 года, как считали те же YFull), к А1 – 62 снипа (8900 лет), возраст А0 не выставлен, но для А1 выставлено 146300 лет «возраста». Опять загадка, как это может быть.
Судя по этим данным, на протяжении 100 тысяч лет между А00 и А0-Т, гаплогрупп между ними вовсе не образовывалось, и к чему те 62 снипа относятся – непонятно. Но здесь важнее то, что А0 – это «африканская» гаплогруппа (опять по нынешнему месту жительства), а А1 – неафриканская. На схеме выше показано семь субкладов гаплогрупы А0, и снипов никаких из них у нас, читателей Переформата, нет, а снипы гаплогруппы А1 есть у нас всех (я не буду больше повторять, что это относится к обладателям Y-хромосомы, то есть к мужчинам, и что «читатель Переформата» относится к неафриканцам, потомкам гаплогруппы ВТ, которые не перебрались на жительство в Африку).
Повторю, что «африканских» снипов А0 у нас нет, можно не искать, нет ни одного из 899 тех снипов. Не выходили наши предки из Африки, или, во всяком случае, не являются потомками ни А00, ни А0.
Надо сказать, что в Африке носителей гаплогруппы А0 обнаружено немного. Scozzari et al (2012) указывали, что только после долгих поисков нашли трех (!) носителей гаплогруппы А0, все в Камеруне, и отмечали, что эта гаплогруппа имеет в Африке очень низкую частоту встречаемости.
Ниже приведен список из 62 снипов гаплогруппы А1, из которых у меня есть L986, L989, L1002, L1009, L1112, V238, V241, V250. Остальных нет потому, что не все снипы у каждого определяются.
Третья вилка – расхождение А1 на A1a и A1b. Потомки A1a сейчас живут в Африке и по всему миру (в частности, в ряде европейских стран и на Ближнем Востоке). Снипы А1b есть у всех нас, читатели Переформата. Но это опять не значит, что наши предки вышли из Африки. Это опять промежуточные снипы, и никого с терминальными снипами А1b пока не выявлено, ни в Африке, ни вне Африки. Поэтому называть его «африканским» опять никакого смысла не имеет. По данным, приведенным YFull, к A1a ведет 495 снипов (это 71 тысяча лет, если снип образуется в среднем раз в 144 года), к А1b – 44 снипа (6300 лет), но почему-то им опять было выставлено по 132600 лет «возраста». Опять загадка, на каком основании так делается.
Хотя носители гаплогруппы A1a разбросаны по всему миру (поэтому в любом случае это не «африканская» гаплогруппа, даже по такому условному критерию, как современное место жительства), их мало. И.Р. Рожанский уже приводил в своей статье «Африканцы на севере Европы. Африканцы ли?», что гаплогруппа A1a-M31 чрезвычайно редкая, и даже в Африке ее наличие исчисляется в лучшем случае 4.4% (в Западной Африке – Гвинея-Бисау, Гамбия, Сенегал, Мали, Нигер, Марокко), и 0.4% среди афроамериканцев. Этому же вторит статья Scozzari et al. (2012), что им удалось найти в Африке для геномных иследований только двоих (!) носителей A1a-M31.
Ниже приведен список из 44 снипов гаплогруппы А1b, но в той системе, которую использует YFull. В классификации ISOGG в гаплогруппе A1b насчитывается 47 снипов, и ни один не пересекается по индексам с приведенными ниже. Из этих 47 снипов у меня есть L989, L1009, L1013, L1053, P108, V221. Остальных нет, потому что, как указывалось выше, не все снипы у каждого определяются.
И вот здесь снипы гаплогруппы A1b приоткрывают загадку. Воспроизведем еще раз фрагмент лесенки снипов гаплогруппы A1:
Мы видим, что от нее вилкой отходят субклады A1a и A1b. Мы уже узнали, что А1а встречается в Европе, на Ближнем Востоке, в Африке, причем нет никаких данных, что он изначально был африканским. Субклад A1b встречается у всех неафриканцев как промежуточный, в цепочке снипов. Субкладов, нижестоящих от A1b, а это A1b1, A1b1a, A1b1a1, A1b1a1a, A1b1a1a1, A1b1a1a2, A1b1a1a2a, A1b1a1a2b – у неафриканцев нет, возможно, потому, что параллельно с A1b1 образовался субклад BT (по даным YFull примерно 126 тысяч лет назад), и A1b1 ушли в Африку, а ВТ остался вне Африки. То есть у неафриканцев имется основная линия A1b > BT, и потомки ее составляют все гаплогруппы от В до Т. И вдруг выясняется, что нижестоящая линия, в субкладе A1b1a1- M23, P3, обнаружилась в Южной Африке среди носителей койсанских языков. Но это – довольно далекий по цепочке вниз субклад, который, по-видимому, отошел от неафриканцев в древние времена. Иначе говоря, не неафриканцы отошли от африканцев, а наоборот, африканцы отошли от неафриканцев, мигрировав в Африку. Опять – не выходили наши предки из Африки. Было наоборот: современные африканцы – потомки неафриканцев.
Четвертая вилка – расхождение A1b на A1b1 и BT. A1b1 и нисходящие субклады считаются африканскими – по нынешнему месту проживания его носителей, хотя родительская гаплогруппа A1b (как промежуточный субклад в цепочке снипов) насчитывает не менее 2-3 миллиардов неафриканцев, а нижеследующие субклады от A1b1 и ниже – обнаружены всего у нескольких африканцев. Вот и считайте, кто от кого произошел. Ответ ясен – африканцы от неафриканцев. Повторю – не выходили наши предки из Африки.
Итак, потомки A1b1 сейчас живут в минимальном количестве в Африке, а потомки гаплогруппы ВТ – это все неафриканские гаплогруппы от В до Т. По данным YFull, к A1b1 ведут всего 26 снипов, к субкладу A1b1a ведут всего 2 снипа, к A1b1a1 – 11 снипов, ни одного из них у нас нет. Датировок ни к одному из нисходящих от A1b1 субкладов в списке YFull нет, и понятно, почему – их носители крайне редко встречаются, да и данных нет. Снипы ВТ есть у всех нас, читатели Переформата.
К сводной гаплогруппе ВТ ведут 403 снипа, и многие из них есть у нас. У гаплогрупп категории А их, естественно, нет. Датировка ВТ по данным YFull – примерно 126 тысяч лет, хотя 403 снипа – это примерно 58 тысяч лет. Некоторые могут подумать, что эти снипы – от родительской гаплогруппы A1b, но та датирована той же компанией YFull примерно 133 тысячами лет назад, то есть всего на 7000 лет раньше. Опять остается неясным, к чему те 403 снипа относятся, и откуда тянутся. Самая загадка в том, как «параллельные» гаплогруппы A1b1 и BT, от одного общего предка A1b, и к которым идут 26 и 403 снипа, соответственно, могут оказаться с одинаковой датировкой – 126 тысяч лет.
Ниже приведен частичный список снипов сводной гаплогруппы ВТ. Как уже отмечалось, многие из них есть у каждого неафриканского мужчины. У меня есть, например, L418, L604, L962, L970, L977, PF1408 и другие.
Итак, первая часть этой статьи безоговорочно показывает, что «африканские ДНК-линии» совсем не древние, в противовес тому, чем нас пичкают популяционные генетики уже 30 лет. Если вести «древность» от общих предков с приматами, то африканские линии ничем не отличаются от неафриканских, имеют одну и ту же глубину. Если считать от времени жизни общих предков современных мужских популяций – по племенам или по территориям, то африканские ДНК-линии весьма мелкие, намного мельче, чем неафриканские. Если считать от образования гаплогрупп, то никто не знает, на каких территориях или континентах они образовались, поэтому присваивать им названия «африканские» – некорректно. Более того, носителей этих гаплогрупп, категории А, в Африке очень мало. К этому могут быть разные причины – прошли бутылочные горлышки популяции, вымерли, или прибыли в Африку относительно недавно – большого значения не имеет, они не «древние», в том смысле, который вкладывают попгенетики. В итоге имеем то, что изображено на заглавной иллюстрации поста.
Так что с этим разделом закончили – нет в Африке «древних ДНК-линий». Но есть такое понятие, как совокупные ДНК-линии. Это тогда, когда сами ДНК-линии мелкие, но они – ошметки древних раскидистых деревьев гаплогрупп и субкладов. Мы видим только верхушки, но экстраполяция верхушек, в данном случае гаплотипов этих «ошметок», позволяет увидеть, когда жили общие предки этих разбросанных гаплотипов. Мы уже видели это выше – сравнение гаплотипов «мелких» популяций гаплогрупп А00 и А0 показывает, что они расходятся на астрономические расстояния. Сами «ошметки» имеют общих предков всего несколько сотен лет назад, но они расходятся на полмилиона лет, и отсюда следует, что их общие предки жили около 250 тысяч лет назад. В этом отношении африканские ДНК-линии отличаются от неафриканских. Рассмотрим это подробнее, в этом упрятана одна из основных загадок человечества.
Загадка была впервые сформулирована в 2012 году, в статье (Klyosov, Rozhanskii, Advances in Anthropology, 2, No. 2, 80-86). Суть ее состояла в том, что все ДНК-линии современных гаплогрупп от В до Т, то есть неафриканских гаплогрупп, сходятся к общему предку, который жил 64000±6000 лет назад, хотя разрозненные линии африканских (по нынешнему месту жительства) гаплогрупп сходятся к общему предку 132 тысяч лет назад (хотя сами разрозненные линии датируются временами жизни общих предков всего лишь 5500, 5000 и 600 лет назад). При этом все неафриканские линии (включая гаплогруппу В) отстоят исключительно далеко от африканских линий, ясно показывая, что неафриканские линии не могли произойти от африканских. Например, базовые гаплотипы гаплогрупп А0 и В отстоят друг от друга на 270 тысяч лет (между ними 18 мутаций в «медленных» 22-маркерных гаплотипах; для сравнения, базовые гаплотипы R1a и R1b отстоят друг от друга всего на 8 мутаций, то есть примерно на 54 тысячи лет, и это помещает их общего предка на 27 тысяч лет назад), иллюстрация увеличивается по клику.
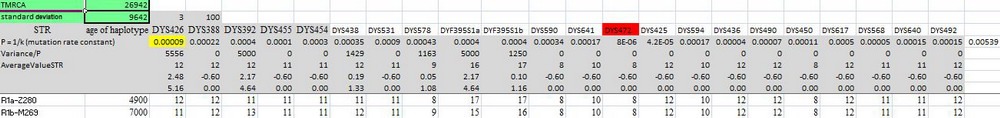
Ясно, что если между гаплогруппами А0 и В 270 тысяч лет, то вторая никак не могла образоваться от первой в течение всего периода существования Homo sapiens, не говоря о 70 тысячах лет назад, которые попгенетики отводят для времени образования гаплогруппы В из А. Это аналогично примеру выше для R1a и R1b, в котором 54 тысячи лет расстояния между ними никак не могут допустить, что одна образовалась из другой; они обе образовались от одного общего предка, которым в данном случае являлась гаплогруппа R1.
Гаплогруппа А0, далеко отстоящая от гаплогруппы В, здесь не исключение. Подобные огромные расстояния отделяют гаплогруппу В от любых гаплотипов в субкладах категории А, например, 250 тысяч лет от A0a-L981, 280 тысяч лет от A1b-M32, 310 тысяч лет от европейского кластера гаплогруппы A1a-M31, 360 тысяч лет от другого кластера той же гаплогруппы, 320 тысяч лет от арабского кластера той же гаплогруппы, 350 тысяч лет от A0a-V150, 356 тысяч лет от A0a-L979, 346 тысяч лет от A0a1a-V151, и так далее. Самое большое расстояние отделяет гаплогруппу В от А00 – 450 тысяч лет, то есть их общий предок жил примерно 225 тысяч лет назад, на утренней заре Homo sapiens.
Такая же картина наблюдается и для гаплотипов всех гаплогрупп от В до Т. Например, растояние от гаплогруппы С до А0-L896 составляет 357 тысяч лет, до A0a1-V150 – 300 тысяч лет, и так далее. Но расстояние от В до С составляет всего 120 тысяч лет, то есть их общий предок жил примерно 60 тысяч лет назад. Это находится в полном соответствии с картиной, что гаплогруппа В образует единый кластер с гаплогруппами от В до Т, и не является по происхождению «африканской» гаплогруппой.
Видимо, уже всем ясно, что гаплогруппа В не образовалась из какой-либо гаплогруппы категории А (поскольку нет гаплогруппы А как таковой, есть А00, А0-Т, А0, А1, А1a, A1b и так далее), а все они ведут свое начало от общего предка. Этот общий предок жил примерно 160 тысяч лет назад, как показано в упомянутой выше статье (Klyosov, Rozhanskii, Advances in Anthropology, 2, No. 2, 80-86). Если исходить из датировок, приведенных на сайте YFull, то основатели гаплогрупп А00 и A0-T жили раньше этого времени, причем гаплогруппа А00 осталась в известном смысле тупиковой, нисходящие от нее субклады и прочие ДНК-линии неизвестны. A0-T прошла своими снипами как к африканцам, так и к неафриканцам, как показано выше, поскольку образовалась еще до разделения на африканскую и неафриканскую ветви. Разделение на эти ветви соответствует временам непосредственно до образования гаплогрупп А0 и А1, причем А0 ушла в Африку, А1 осталась вне Африки. Впоследствии носители многих гаплогрупп мигрировали в Африку, например, A1a и нижестоящие субклады, A1b1 и нижестоящие субклады, поэтому их снипов у нас нет, как описано выше.
Все это описано в целом в упомянутой выше статье (Klyosov, Rozhanskii, Advances in Anthropology, 2012), которая, кстати, набрала рекордное количество просмотров (около 90 тысяч) и скачиваний (около 20 тысяч), но академическое сообщество предпочло ее не заметить. Слишком многое она меняет в представлениях. Основная иллюстрация в той статье следующая:
Номенклатура в этой иллюстрации несколько устаревшая, в ней нет гаплогруппы А00, которая в 2012 году была неизвестной, но суть ее остается правильной. Гаплогруппа альфа – это условная гаплогруппа, от которой пошло разделение эволюционного Y-хромосомного дерева человечества на африканскую (слева) и неафриканскую линии. В настоящее время все гаплогруппы от В до Т, то есть гаплогруппы неафриканского происхождения, сходятся к условной же бета-гаплогруппе, которой на самом деле является сводная гаплогруппа ВТ. Загадка – куда делась связка между альфа- и бета-гаплогруппой, протяженностью в сто тысяч лет? И, кстати, куда делись африканские ДНК-линии, которые на левой части диаграммы полностью экстраполированы, от общих предков всего от тысячи до нескольких тысяч лет давности, вглубь времен?
Со времени публикации той статьи прошло уже более трех лет, в базах данных и в Проектах с тех пор накопилось много гаплотипов, в том числе протяженных, 111-маркерных, и появилась возможность проверить количественные выводы статьи. То, что расхождение африканской и неафриканской ДНК-линий произошло примерно 160 тысяч лет назад, мы уже проверили. Перейдем ко времени жизни общего предка всех неафриканских гаплогрупп – от В до Т. Ниже – дерево гаплотипов этих гаплогрупп. Оно построено по 111-маркерным гаплотипам, и сами гаплогруппы показаны по окружности дерева. Для каждой из 18 гаплогрупп использовали 10-25 гаплотипов, взятых произвольно из выборки, всего 403 гаплотипа nd 111-маркерном формате. Гаплогруппы категории А на дереве не оставили, так как они явно «вылезали» из кругового дерева, поскольку, как мы уже показали, они удалены по мутациям от всех других гаплогрупп.
Видно, что все гаплогруппы от В до Т образуют достаточно единую систему ДНК-линий (выраженную здесь через 111-маркерные гаплотипы), так что про некую «африканскую гаплогруппу В» говорить не приходится. Она такая же «автохтонная африканская», как R1b – «автохтонная европейская». Это застарелая болезнь попгенетиков – то, что видят сейчас, значит, так было всегда. Болезнь называется «отсутствие научного подхода», или «отсутствие научной школы», что, впрочем, одно и то же, а если по-русски, то «думать не умеют».
403 гаплотипа в 111-маркерном формате, то есть почти 45 тысяч аллелей – это крепкий орешек, если считать вручную. Более того, за десятки тысяч лет в них накапливаются десятки тысяч мутаций, помимо того, огромное количество возвратных мутаций, что делает 111-маркерную систему такой древности неустойчивой, и неприемлемой для расчетов, поскольку датировки значительно занижаются. Поэтому для данных случаев разработана 22-маркерная панель из самых «медленных» маркеров. Они настолько устойчивы во времени, что для всех 403 гаплотипов за десятки тысяч лет маркер DYS472 вообще не мутировал ни разу, DYS436 – всего 30 раз, DYS590 – 42 раза, и так далее. Сравните – маркер DYS390 за это же время должен мутировать 2270 раз, то есть в среднем 5.6 раз на гаплотип. При таких частотах мутации расчет вклада обратных мутаций уже практически невозможен, система нестабильна.
А вот «медленная» 22-маркерная панель с малыми скоростями мутаций позволяет проводить расчеты на десятки и сотни тысяч лет вглубь, и даже на миллионы лет, как было показано выше на примере расчета времени жизни общих предков человека и шимпанзе. Более того, разработанный калькулятор Килина-Клёсова проводит расчеты по каждому маркеру отдельно, используя квадратичный метод, и поэтому не требует введения поправок на возвратные мутации. В итоге ДНК-генеалогия получила незаменимый метод расчета массивных серий гаплотипов в любом формате и любой древности общего предка (Килин и Клёсов, Вестник Академии ДНК-генеалогии, 2015, т. 8, № 3, 321-375, http://dna-academy.ru/kilin-klyosov/). Расчет дерева из 403 гаплотипов, приведенного выше, показал, что общий предок гаплогрупп от В до Т жил примерно 63 тысячи лет назад (результат без округления – 63036 лет, что, конечно, с такой точностью совершенно излишне). Это почти точно совпадает с рассчитанной ранее (Klyosov and Rozhanskii, 2012) величиной 64±6 тысяч лет.
Теперь осталось подойти к решению сформулированной в той статье загадки – повторим ее опять: куда же делись предки Y-хромосомных линий в Африке ранее 5-10 тысяч лет назад, и у неафриканцев ранее 64±6 тысяч лет назад? Причем у африканских линий группы А можно косвенным путем, а именно при сравнении протяженных гаплотипов с «неглубокими» общими предками, достичь времен жизни общих предков 132 тысячи лет назад, а у неафриканцев, как их гаплотипы ни комбинировать и ни сравнивать, глубже 64±6 тысяч лет уйти не получается. Более древних общих предков как корова языком слизнула. Что же это за корова была такая?
Некоторые предположения в цитированной выше статье сделаны были – упомянуто возможное последствие извержения вулкана Тоба в Индонезии, около 70 тысяч лет назад. Не исключено было резкое похолодание, возможно, как результат извержения того же вулкана, причем, по мнению специалистов, ранг катастрофичности глобального похолодания намного выше, чем извержения вулкана, но это все оставалось довольно абстрактными предположениями. И вообще – чтобы извержение вулкана в Индонезии привело к гибели почти всех людей на планете – это кажется маловероятным, но все это «по понятиям». Нужно было мнение специалиста, пусть мнение и спорное, неоднозначное, но главное – подтвержденное конкретными данными.
И такой специалист по данной проблеме есть. Знакомьтесь – Валерий Павлович Юрковец, профессиональный геофизик, палеоклиматолог, специалист в области влияния климатических катастроф, в том числе импактного характера, на миграции древнего человека, член Академии ДНК-генеалогии с начала ее образования несколько лет назад.
Продолжение следует…
Анатолий А. Клёсов,
доктор химических наук, профессор
Осталось подойти к решению сформулированной в статье (Klyosov, Rozhanskii, 2012) загадки – повторим ее опять: куда же делись предки Y-хромосомных линий в Африке ранее 5-10 тысяч лет назад, и у неафриканцев ранее 64±6 тысяч лет назад? Причем у африканских линий группы А можно косвенным путем, а именно при сравнении протяженных гаплотипов с «неглубокими» общими предками, достичь времен жизни общих предков 132 тысячи лет назад, а у неафриканцев, как их гаплотипы ни комбинировать и ни сравнивать, глубже 64±6 тысяч лет уйти не получается. Более древних общих предков как корова языком слизнула. Что же это за корова была такая?
Некоторые предположения в цитированной статье сделаны были – упомянуто возможное последствие извержения вулкана Тоба в Индонезии, около 70 тысяч лет назад. Не исключено было резкое похолодание, возможно, как результат извержения того же вулкана, причем, по мнению специалистов, ранг катастрофичности глобального похолодания намного выше, чем извержения вулкана, но это все оставалось довольно абстрактными предположениями. И вообще – чтобы извержение вулкана в Индонезии привело к гибели почти всех людей на планете – это кажется маловероятным, но все это «по понятиям». Нужно было мнение специалиста, пусть мнение и спорное, неоднозначное, но главное – подтвержденное конкретными данными. И такой специалист по данной проблеме есть. Знакомьтесь – Валерий Павлович Юрковец, профессиональный геофизик, палеоклиматолог, специалист в области влияния климатических катастроф, в том числе импактного характера, на миграции древнего человека, член Академии ДНК-генеалогии с начала ее образования несколько лет назад. В.П. Юрковец написал статью в сентябрьский выпуск Вестника Академии ДНК-генеалогии, и я здесь кратко перескажу ее основные результаты и выводы, с моими дополнениями по ДНК-генеалогии.
В.П. Юрковец приводит убедительные доказательства того, что между 57 и 71 тысяч лет назад крупное космическое тело упало в Тихий океан, что, в свою очередь, вызвало мегацунами, так называемое космогенное мегацунами. Размер этого тела был, вероятно, сопоставим с Ладожским импактом – падением на север Русской равнины массивного астероида около 11 километров в поперечнике, но по катастрофическим последствиям превосходящим его на порядки. Как пишет В.П. Юрковец, каменный метеорит диаметром 100 м вызывает на суше разрушения в радиусе немногим более километра, что моментально приводит к возникновению очень сильного цунами сейсмического происхождения. Но при падении крупного космического тела в океан разрушительные последствия неизмеримо более сильные. При этом возникают несколько следующих друг за другом гигантских волн.
Первая – от непосредственного удара о водную поверхность, вторая – от схлопывания (заполнения водой) сформированного ударом водного кратера (каверны), третья – от кратерообразования в океанском дне, то есть в толще пород, слагающих дно океана, и от сопутствующих вторичных процессов. При диаметре метеора 11 км он превосходит по размерам глубину океана практически в любом месте, достигает океанского дна в момент удара, и без труда может пробить тонкую (средняя толщина 7 км) океаническую кору, что приведет к извержению и возникновению дополнительной волны цунами. В открытом океане цунами распространяются со скоростью, достигающей 700 км/час, то есть скорости самолета. Космогенные мегацунами образуют волны, достигающие в высоту нескольких километров, и перехлестывающие самые высокие горные хребты. Это не выдержки из фантастических произведений, это описание того, что на Земле уже происходило, и к тому есть доказательства, в виде многочисленных фотографий последствий этих явлений и соответствующих расчетов, во множестве приведенных в статье В.П. Юрковца. Мегацунами – вот что «как корова языком слизнула» человечество в середине 70-го тысячелетия до нашей эры, 64±6 тысяч лет назад, как показывает ДНК-генеалогия, что совпадает с 4-й морской изотопной стадией, или MIS 4 (71-57 тыс. лет назад), выделяемой палеоклиматологами по резкому ухудшению климатических условий.
Но там было не только резкое ухудшение климатических условий, ухудшение может быть по разным причинам. И извержение вулкана Тоба около 70 тысяч лет назад может случайно (или не случайно) совпасть по времени с исчезновением почти всего человечества. То, что именно в то время на палеоклиматических кривых, приведенных в статье В.П. Юрковца, наблюдается резкое увеличение пыли в атмосфере, понижение температуры, увеличение массы ледников – это, конечно, человечеству не подарок, но о мегацунами пока не говорит. А вот тот факт, что в дополнение ко всему этому палеоклиматические кривые свидетельствуют о резком засолении всей Антарктиды, то есть захлестывании всего материка морской водой, это уже аномалия глобального характера. Тем более что ледовые керны на обоих полушариях показывают резкое увеличения натрия во льду, который мог поступить только с морской водой. Это более чем экстраординарное событие произошло 68 тысяч лет назад. Таким образом, как подчеркивает В.П. Юрковец, все катастрофические события того времени оказались связаны в единый узел.
Еще о катастрофичности тех событий. Судя по размерам кальдеры Тоба (около 100 километров по длинной оси), космический толчок, спровоцировавший его извержение, не уступал по мощности Ладожскому событию, извержение которого сформировало собственную кальдеру оседания около 130 километров в диаметре и спровоцировало извержения вулканов, как минимум на Кавказе и в Италии. Как уже отмечалось, Ладожский астероид имел в поперечнике около 11 километров (Юрковец, 2012), и космическое тело, породившее мегацунами в Тихом океане, вероятно, не уступало ему в размерах. По крайней мере, вулканические последствия двух падений сопоставимы. Диаметр Ладожской астроблемы составляет больше 100 километров. При диаметре астероида, превышающем глубину океана, слой воды не оказывает существенного влияние на формирование кратера, следовательно, размер кратера, образованный падением астероида, вызвавшего мегацунами в Тихом океане, не может быть меньше нескольких десятков километров в поперечнике. Это уже величина, вполне различимая на картах океанического дна Google, и об этом речь пойдет ниже.
Космогенное мегацунами оставило неизгладимые следы на континентах в виде гигантских знаков ряби, называемых у специалистов «гигантская рябь течения», а также «ламинарных супертечений» и «турбулентных сверхпотоков». Я приведу здесь лишь несколько характерных фотографий из статьи В.П. Юрковца, у него их там много десятков. Существует теория дилювиального морфолитогенеза (разработанная доктором географических наук А.Н. Рудым), для гигантских потоков, вызванных прорывами ледниково-подпрудных озёр, и она иллюстрируется на активных русловых формах рельефа высотой от 2 до 20 метров. Это, конечно, ни в какое сравнение не идет с рельефами, вызванными мегацунами 57-71 тыс. лет назад на всех континентах планеты, как показано ниже.
Гигантские знаки ряби течения – это система вытянутых гряд или цепи дюн серповидной формы, синхронно ориентированных навстречу образовавшему их потоку, иногда разделенных перемычками, сопоставимых с высотами основных дюн. Специалистам известны и углы образования проксимальных и дистальных склонов гряд ряби течения и дюн, их профилей, пригребневых частей, и так далее. По ним на аэро- и космоснимках определяют направления дилювиальных потоков прошлого. Типичные размеры гигантских знаков ряби русловых форм рельефа – высота от 2 до 20 м при длине волны (растояние между цепями дюн) от 5-10 м до 300 м. Вот, например, аэрофотоснимок классической гигантской ряби течения в Курайской впадине на Алтае (Рудой, 2006), приведенный В.П. Юрковцом в своей статье:
А вот уже следы космогенного мегацунами, на многочисленных примерах, исследованных В.П. Юрковцом на разных континентах. Это – пустыня Намиб в юго-западной Африке. Те же характерные морфологические признаки, те же гряды, те же перемычки, то же соотношение высоты дюн и максимальной длины волны – только на порядок выше, чем для гигантской ряби течения более традиционных «мегапотоков». В пустыне Намиб высота дюн – до 200 м, длины волн – до 3 километров. Средние величины – 150 м и 2.2 километра, что опять повторяет «классическое соотношение». Здесь и далее – Google maps, за исключением наземных фотографий.
Следы космогенного мегацунами – гигантская рябь течения в пустыне Намиб в юго-западной Африке (из статьи В.П. Юрковца). При увеличении картина выглядит еще более впечатляюще – протяженность дюн может достигать 20 километров:
Не нужно здесь обманываться словом «дюны», это не песчаные дюны, они только присыпаны песком. Основа дюн – грубообломочный (дилювиальный) материал, крупный щебень, только мегаразмера, опять мега, других измерений не подобрать.
Вот так дюны выглядят:
Ориентация дистальных склонов дюн – на восток, то есть поток пришел со стороны моря. Мощные водные потоки выработали много характерных узких каньонов с многочисленными следами кавитации на стенах:
Кавитация – это следы взрывного схлопывания каверн, образующихся при перепадах давления в жидкости, движущейся с переменной скоростью. Вследствие свойства несжимаемости, фронт ударной волны, образующийся при схлопывании каверны, имеет большую крутизну и подобен взрыву. Поэтому кавитация обладает большой разрушительной силой, она оставляет особые следы на материале, на который воздействовал скоростной поток. К ним относятся многочисленные следы «микровзрывов» – поверхности сколов в дилювии и материале, контактирующим с потоком, движущимся с большой скоростью. Ни один другой агент – ветер, попеременно замерзающая и оттаивающая вода, обычное течение, волно-прибойная деятельность и пр. не оставляет подобных следов.
По расчетам, глубина (высота) потока, накрывшего пустыню Намиб, могла составлять от 9 до 15 километров. Эти и другие расчеты показывают, что следы гигантского цунами, которые присутствуют на всех материках, были оставлены несколькими последовательными волнами, образованными единым падением массивного космического тела в Тихий океан:
Проявления космогенного мегацунами многочисленны. Это, например, вортексы («исполинские котлы»), выработанные гигантскими водоворотами, несущими камни, и достигающие десятков и сотен метров в диаметре, хотя в Северной Африке есть и гигантский вортекс диаметром около 40 километров.
Эрозионно-кавитационные образования – вортексы («исполинские котлы»), выработанные гигантскими водоворотами, несущими камни. Вортекс на фото – диаметром 80 метров, в Австралии. Фото В.К.Гусякова (Алексеев, Гусяков, 2011). Вортекс в Мавритании, Северная Африка:
Это также геоскульптурные эрозионно-кавитационные формы рельефа бассейна реки Колорадо
на восточных склонах Кордильер и Скалистых гор
в высокогорных регионах Анд в Южной Америке
на Дальнем Востоке (Китай), в Сибири (Красноярские столбы), на Урале (плато Маньпупунёр)
Но вернемся к волнам гигантской ряби. Итак, следы космогенного мегацунами на всех континентах показывают, что высота гигантской волны, возникшей в результате падения крупного космического тела в Тихий океан, достигала высоты до 15 км. Она преодолевала высочайшие горные хребты и даже континенты. Встречные волны ударились друг о друга в районе северной Африки, именно там зафиксирована хаотичная картина гигантских знаков ряби, «супертечений», «мегапотоков» и прочих свидетельств движения огромных масс воды. Наиболее близкой к центру мегацунами оказалась Австралия, где оставленная в пустыне Симпсона гигантская рябь течения отличается наиболее строгой геометрией, из всех встречавшихся:
Здесь длина волны гигантской ряби составляет около 300 метров. Протяжённость дюн – до 20 и более километров. И эта единообразная картина наблюдается по всей пустыне, протяженностью более 500 километров. То есть это был ровный, без внутренних флуктуаций и завихрений, единый и невероятно грандиозный поток, который, вероятно, даже «не заметил» Австралию – предыдущая тысяча километров суши, включая среднегорье, по которой он прошёл до пустыни Симпсона, никак не сказались на его «ламинарности».
В Северной Америке мегацунами перехлестнуло Кордильеры и Скалистые горы и фотографии разрушений на противоположной от океана, восточной стороне гор, приведены выше. В пустыне Хила, расположенной в районе американо-мексиканской границы на территории американских штатов Аризона и Калифорния и мексиканского штата Сонора, к северу от Калифорнийского залива, оно оставило гряды гигантских дюн – гигантскую рябь течения с высотой дюн до 150 м и длиной волны, превышающей 4 километра – на километр больше, чем в пустыне Намиб. Морфология дюн такая же в обеих пустынях – в Африке и Северной Америке. Пройдя дальше, в штате Колорадо, суперпоток быстро преодолевал турбулентность при выходе из долины реки Литл Колорадо, и создал рельеф, для которого даже нет названия в существующей терминологии ландшафтов – дюны в виде широких ячеистых полос и переходом в протяженные гребни, разделенные впадинами:
В одном месте по ходу движения поток встретил препятствие – одиночную вершину, и вот к чему это привело:
Здесь физико-математическая реконструкция выявила действие трех ударных суперпотоков, причем первый поток перехлестнул через вершину, высота второй волны была меньшей, третьей волны – еще меньшей, она породила турбулентные потоки, отходящие непосредственно от подножия горы. В итоге долины, отходящие от горы, имеют вихревую природу. Еще одно поле дюн турбулентности находится в Аризоне, его размер по диагонали составляет около 4 километров:
В Южной Америке волнам мегацунами противостояли величайшие горные хребты мира, к тому же параллельные друг другу и поперек удару суперпотока. Но это не стало помехой – поток просто перехлестнул их и к тому же пропилил в самых высокогорных частях Анд целую систему долин-спилвеев и изменил направления рек, которые стали течь тоже поперек хребтов (верховья Рио-Гранде и других рек). Примеры геоскульптурных образований приведены выше, на фотографиях в высокогорных регионах Анд в Южной Америке. Там даже нет окатанного галечника, что свидетельствует о кратковременности потока. Гигантские ряби течения встречаются, например, в Аргентине, на плато Альтиплано, и на равнинных территориях, которые даже не закрываются сплошным лесом. Там средняя длина волны превышает 4 километра, что показывает, что удар мегацунами по Южной Америке был сильнее, чем по Северной Америке.
В Азии мегацунами оставило многочисленные следы в виде гигантской ряби – например, в восточной части пустыне Гоби, со средней длиной волны 3.4 километра:
На западной части пустыни Гоби длина волны достигала уже 3.2 километра, то есть поток несколько ослабевал при продвижении на запад. В Монголии, в долине реки Завхан, длина волны уменьшилась уже до полутора километров:
В районе Каспия картина следов мегацунами стала более хаотичной, но вал с огромной скоростью катился дальше, в Европу, которая, видимо, пострадала меньше всех. Южный рукав мегапотока ударил по Памиру и Гималаям, образовав классическую гигантскую рябь течения в пустыне Такла-Макан, с длиной волны до 3 километров:
При этом отраженные волны мегацунами встретились, что привело к образованию серии «дюн турбулентности», направленную дистальными склонами на северо-восток:
По ходу своего движения поток пропилил, наверное, самый большой в мире спилвей – Алайскую долину, отделившую Памир от Гиссарского и Зеравшанского и Алайского хребтов, по которой вода устремилась в гигантский бассейн Каспийского и Аральского морей, перехлестнул через горы на Иранское плато, и сформировал там сразу несколько спилвеев. Пример – ущелье Танге Ваши, неподалеку от современного Тегерана:
Следы мегапотока в Индии-Пакистане, в пустыне Тар, протяженностью 850 км (северо-запад Индии и юго-восток Пакистана), где зоны турбулентности чередуются с ламинарным течением:
Гигантская рябь течения встречается на юго-востоке Аравийского полуострова, откуда мегаволна ударила по Африке, пройдя континент с юга на север. Пример ламинарного супертечения к западу от низовий Нила:
А вот – и более обширная картина в Африке:
Еще доказательство мегацунами – солёные бессточные высокогорные озера. Самое высокое в мире – соленое бессточное озеро Намцо в Тибете, высота над уровнем моря 4718 метров. В попытках объяснить это явление была выдвинута гипотеза, что соль поступала с дождевыми водами, текущими по горным породам, хотя воды текут везде, но озера почему-то пресные. Бессточное озеро Балхаш (высота над уровнем моря 340 м), например, состоит из двух частей – соленая восточная, и пресная западная, соединенные небольшим проливом. В пресную впадает река, которая несет соли, но это не дает особой солености. Отчего другая часть почти на порядок более соленая – никто не объясняет, и в Википедии кроме описания этого факта, а также что соленость пресной части 0.74 г/л, а соленой части 3.5-6.0 г/л, больше ничего нет. В.П. Юрковец относит солёность Балхаша за счет реликта некоего события, которое с поступлением соли из окрестностей никак не связано. Какое это могло быть событие – примерно уже ясно, хотя бы как вариант.
Наконец, на последствия космогенного мегацунами могут указывать океанские морфологические структуры, образованные, видимо, мощнейшими потоками, которые буквально за минуты и часы вырабатывали глубоководные долины, каменный материал которых тут же сбрасывался в океанический простор. Пример – южный прорыв между Южной Америкой и Антарктидой
Северный прорыв, состоящий из трех шлейфов, возможно, трех волн космогенного мегацунами, между Южной и Северной Америками:
При этом сформировались глубоководные желоба (на снимках выделяются темно-синим, почти черным цветом), находящиеся на фронтальной части дуг (шлейфов), сформированных обломочным материалом, предельные глубины которых составляют между Южной Америкой и Антарктидой 8428 метров, между Америками 8385 метров. Глубже их только впадины, образованные столкновением тектонических плит. Эти желоба были сформированы (вынесены) классическим делювием (тем самым обломочным материалом), который принесен потоком, но не окатан, как показано на фотографии выше.
Полагаю, приведеные здесь данные (они, повторяю, все взяты из статьи В.П. Юрковца в Вестнике Академии ДНК-генеалогии) убедительно свидетельствуют о масштабе космогенной катастрофы, происшедшей на Земле 57-71 тыс. лет назад. Она не могла оставить на планете ничего живого, но, получается, что оставила. Сохранились животные и птицы, хотя известно, что после 50 тысяч лет назад (грубая оценка) большинство животных исчезли с лица Земли. Объяснений нет, разве что «по понятиям», например, что их уничтожил человек. Но сам человек после космогенной катастрофы сохранился в столь небольших количествах, что выживание его висело буквально на волоске.
Рассмотрим сначала неафриканцев. Потомство выжило у минимального их количества, что и привело к эффекту «бутылочного горлышка популяции» 64±6 тысяч лет назад. Фактически, в те времена у неких отца с матерью выжили как минимум двое сыновей, которые тоже дали выжившее потомство, и оно постепенно пошло в количественный рост. Все остальные выжившие после катастрофы люди тоже могли дать потомство, но оно пресеклось, не выжило. В итоге практически все мужчины современности на планете, во всяком случае, те сотни тысяч человек, которые прошли тест на гаплотипы-гаплогруппы Y-хромосомы, сходятся по мутациям в гаплотипах формально к одному общему предку, тому самому патриарху, от которого выжили как минимум двое сыновей, давших выжившее же мужское потомство в середине 70-го тысячелетия назад.
Следует подчеркнуть, что тот патриарх имел в своей Y-хромосоме, разумеется, всю цепочку из сотен тысяч снипов, ведущую от общих предков с приматами, и далее вверх по цепочке снипов Y-хромосомы, через A0-T, A1, A1b, ВТ, как описано в первой части. Носители этих снипов как терминальных погибли в катастрофе, но выживший, ставший патриархом неафриканского человечества, их сохранил. Согласно расчетам компании YFull (http://www.yfull.com/tree/), гаплогруппа ВТ образовалась примерно 126 тысяч лет назад (доверительный интервал 121-132 тыс. лет назад), то есть задолго до катастрофы. Возможно, расчеты потом уточнятся, но примем их за основу, других всё равно нет. Так вот, выживший патриарх имел именно эту гаплогруппу, ВТ. Наверняка сотни тысяч и миллионы погибших в катастрофе имели многие другие гаплогруппы и субклады, образовавшиеся за 60 тысяч лет от образования BT до времени катастрофы, но они погибли, и те гаплогруппы и субклады не были унаследованы. Поэтому отсчет выживших поколений пошел именно от гаплогруппы ВТ, и не от времени ее образования, а от времени бутылочного горлышка, вызванного катастрофой. Эта гаплогруппа и была названа бета-гаплогруппой, по факту выживания, а не по факту образования за 60 тысяч лет до катастрофы.
В отношении женщин пока неизвестно, возможно, их после катастрофы по каким-то причинам осталось намного больше, только толку в том для будущих поколений было немного, если мужчины почти все погибли. Но не исключено, что мужчины с той же Y-хромосомой, то есть от одного отца, родились от многих разных матерей, как и их дочери, которые Y-хромосому не имеют. В таком случае общий предок мтДНК, то есть праматерь, может оказаться значительно древнее, чем 64±6 тысяч лет назад. Но подобные расчеты мне неизвестны, да и методы расчетов по мтДНК не столь совершенны, и продолжают дебатироваться. Тем не менее, в недавней статье (Groucutt et al., Evolutionary Anhropology, 2015) сообщается, что все мтДНК современных неафриканцев сходятся к общему предку, жившему (точнее, жившей) в интервале 60-79 тысяч лет назад. По другим источникам – 60-70 тысяч лет назад (ссылка).
Это – гаплогруппа L3. На схеме ниже (Behar et al., 2012) видно, что старейшая гаплогруппа L0, которую считают «африканской», и которая представляет большую серию из полусотни «африканских» гаплогрупп, найденных в основном к югу от Сахары – среди койсанского населения Южной Африки, а также в Эфиопии, Танзании, Мозамбике и среди пигмеев – не является предковой по отношению ко всем остальным гаплогруппам мтДНК. Более того, время жизни общего предка мтДНК практически совпадает со временем жизни Y-гаплогруппы BT и с временем описываемой здесь космогенной катастрофы.
Наконец, ископаемые неандертальцы в Азии и на Ближнем Востоке тоже датируются не ранее 50-70 тысяч лет (Groucutt et al., 2015), причем сообщается, что все те, для кого были тестированы ДНК, имеют общего предка в том же временном интервале. Иначе говоря, их «разнообразие» было минимальным, все – достаточно близкие родственники.
Возвращаемся к мужской гаплогруппе В. Непонятно, почему в списке YFull время образования гаплогруппы B указано как 83900 лет назад, как и время образования «параллельной» СТ (с доверительными интервалами у обеих от 79700 до 88100 лет назад). С одной стороны, они могли образоваться еще до катастрофы, от той, ранней гаплогруппы ВТ, и уйти, скажем, в Африку, где и выжили, и дали свое потомство. Но гаплогруппа ВТ у «автохтонных африканцев» не обнаружена, да и СТ к Африке никак не относится, тем более с нисходящими гаплогруппами С и F (обе по данным YFull образовались примерно 65 тысяч лет назад, с доверительным интервалом между 62 и 68 тысяч лет), и у каждой много нисходящих снипов, явно не африканских. Да и дерево гаплотипов человечества, приведенное в первой части этой статьи, показывает, что гаплогруппа В образует единую систему с неафриканскими гаплогруппами.
Вывод – что-то с датировкам В и СТ у YFull не то. Они не могли быть ранее 64±6 тысяч лет назад. Правда, взгляд на количество снипов, по которым предположительно YFull должны были вести расчеты, объясняет, в чем у YFull проблемы. Для одинаковой датировки гаплогрупп В и СТ у них должно быть одинаковое число снипов, а там стоит 30 снипов у гаплогруппы В, и 316 снипов у гаплогруппы СТ. Иначе говоря, по данным компании YFull (144 лет на снип), гаплогруппа В должна отстоять от ВТ на 4300 лет, а СТ – на 45 тысяч лет. Это с расчетами YFull с никакой стороны не сходится, в том числе и с постулатом равного количества лет для «параллельных» снипов. Поэтому вопрос пока закроем до пересчетов компанией YFull своих (точнее, чужих) данных.
Итак, с гаплогруппами, и вообще с ДНК-генеалогией человечества, в целом ясно, как и с возрождением его, человечества, после космогенной катастрофы 64±6 тысяч лет назад. Это, правда, ставит другую потрясающую загадку – что же представляло человечество до катастрофы? Какие там были цивилизации? Что они после себя оставили, что не было смыто мегацунами? Понятно, что самих людей не осталось, они были разорваны в клочья ударами волн в смеси с острым щебнем, высотой до 15 км и несущимися со скоростью самолета, перехлестывающих через самые высокие горы мира. Остались только немногочисленные скелетные остатки, с которыми работают палеоантропологи, да и те древние люди в большинстве своем определенно не являются нашими предками, чисто статистически они были предками погибших в космогенной катастрофе. Поэтому сопоставления «архаичности» ископаемых скелетных остатков с антропологией современного человека, скорее всего, являются безотносительными. Это покажет тестирование этих «архаичных» на ДНК, откуда потоком пойдут неизвестные нам гаплотипы и гаплогруппы, во всяком случае после A0-T, А1 и A1b. Но может, и тех не будет.
Излагаемая здесь новая концепция затрагивает и другие загадки, давно волнующие исследователей. Например, загадкой является, как получилось, что предки современных людей совершили столь быстрый переход от каменного века до более современных технологий. Сейчас становится понятным, что люди, выжившие после катастрофы, и выброшенные природой обратно в каменный век, были людьми ментально относительно развитыми, после ста тысяч лет цивилизации, нам, впрочем, пока неизвестной. И свои природные способности они относительно быстро возвращали в ходе тысячелетий выживания после катастрофы, следуя той хрестоматийной экспоненте прогресса человечества.
Еще одна загадка, точнее, комплекс загадок – это находимые время от времени древние объекты и технологии, которые не находят объяснения в рамках существующих представлений, и которые, более того, не могут быть воспроизведены. Это и способы перемещения гигантских каменных блоков, и изготовления блоков пирамид с помощью предполагаемых технологий древности, как, впрочем, и укладка самых древних пирамид, встык, без зазоров на гладкой поверхности, из огромных многотонных блоков, и древнейшие пирамиды из странных материалов, нам сейчас неизвестных, и многое другое. Обычно люди, которые пытаются обсуждать это вопросы, подвергаются осмеянию людишками, к решению сложных вопросов вообще неспособными. Но так устроен современный мир, в котором подавляющее большинство критиков и отвергающих новые выдвигаемые концепции, причем отвергающие «по понятиям», являются мизерными посредственностями. «Фрик» – любимое их словечко.
Перейдем теперь к африканской части человечества. Впрочем, всё, что сказано выше о цивилизациях во времена до космогенной катастрофы, относится и к африканцам, если они там жили до 60 тысяч лет назад. Мы ведь и этого не знаем. Времена общих предков современных «автохтонных» африканцев не превышают 5-10 тысяч лет назад (см. первую часть), а цепочки их снипов, уходящие на сотни тысяч лет назад, не привязаны к Африке. Тем не менее, будем исходить из того, что люди в Африке жили в период до катастрофы, ведь находят же там древние скелетные останки людей с датировками до 200 тысяч лет назад. Мы, повторяю, не знаем, были ли они нашими предками, и не узнаем, пока их кости не будут тестированы на ДНК. Так вот, подавляющее большинство тех людей тоже погибли в космогенной катастрофе, но в отличие от неафриканцев выжили люди, возможно, всего несколько человек, но из разных ДНК-линий. Неафриканец-патриарх, напоминаю, был гаплогруппы ВТ. Патриархи-африканцы были нескольких линий, но мы не знаем, какие они были, где те линии/гаплогруппы образовались, возможно, вне Африки, и их носители пришли туда позже, но до катастрофы. А может, и после катастрофы.
Поскольку крупное космическое тело упало в Тихий океан, как показано на карте-схеме выше, и самым сильным волновым ударам подверглись обе Америки, Азия и Австралия, то Африка оказалось несколько в стороне и на большем расстоянии. Разрушения и там были гигантскими, но какое-то число носителей этих пока неизвестных нам гаплогрупп выжили, вместе с женщинами, носителями наследственных мтДНК. Поэтому мужчины начали свое возрождение не с одной гаплогруппы, а с нескольких. Там самым они получили некоторое «разнообразие», которое впоследствии многократно приумножилось за счет многочисленных миграций в Африку – особенно прибывших гаплогрупп В, E1b, R1b (в Камеруне-Чаде и Южной Африке, потомков буров), J1 и J2 (включая племя Лемба в Южной Африке и им подобных), и так далее. Так в Африке сформировалось «генетическое разнообразие», которое к «автохтонным» африканцам практически не относится. Африка – открытая система, а понятие «разнообразие» к открытым системам не относится.
Хочется верить, что в черной Африке тоже были продвинутые древние цивилизации, но пока свидетельств мало, кроме тех, что на севере Африки, в частности, древнейшие пирамиды и Сфинкс. На них ясно видны кавитационные разрушения и знаки водной эрозии на большой высоте, которые традиционно приписывают действию ветра и песка. В.П. Юрковец придерживается другой точки зрения, и не без оснований. Во всяком случае, его точка зрения должна быть рассмотрена на равных, по гамбургскому счету. Однако про критиков было выше.
Анатолий А. Клёсов, доктор химических наук, профессор