ГЕНЕТИЧЕСКИЕ СЛЕДЫ ИСТОРИЧЕСКИХ И ДОИСТОРИЧЕСКИХ МИГРАЦИЙ: КОНТИНЕНТЫ, РЕГИОНЫ, НАРОДЫ
Е.В. Балановская, О.П. Балановский
Медико-генетический научный центр РАМН, Москва, Россия
Современная геногеография, изучая народы мира по «однородительским» ДНК маркерам (Y-хромосоме, наследующейся по отцовской линии, и митохондриальной ДНК - по материнской линии), реконструирует цепочку последовательно происходивших мутаций («от Адама» или же «от Евы»), определяет место и время их возникновения. Это позволяет проследить процесс расселения человечества по планете. Рассмотрены генетические реконструкции событий истории популяций в диапазоне от 140 тыс. лет до 140 лет. Южная Африка: 140 тыс. лет назад произошло разделение современного человечества на два крупных ствола: бушмены и все остальное человечество. Причем почти все разнообразие мтДНК приходится на народы Африки: лишь малая часть ветвей дала побеги на остальных континентах. Центральная Африка: 70 тыс. лет назад произошло расхождение генофондов самого низкорослого и самого высокорослого населения планеты -пигмеев и бантуязычных народов. Неолит Европы: анализ ДНК древних популяций и современной изменчивости генофонда показал, что, хотя распространение земледелия внутри Европы носило характер «культурной диффузии», его появление в Европе связано с миграцией первых земледельцев из Передней Азии. Финикийцы: прослежено их расселение по Средиземноморью. Крестоносцы: выявлен их генетический след в Леванте. Киргизы: дальняя миграция из енисейских степей на высокогорья Памира. Русский народ: мощный генетический пласт восточных славян и ассимилированных финно-угорских племен и отсутствие генетических следов «монгольского ига». Русский Север: огромные различия между северными популяциями и обедненность спектра ДНК гаплогрупп из-за дрейфа генов. Казаки Северного Кавказа: терские казаки включили в свой генофонд значительное число Y- хромосом от коренного населения Кавказа, а кубанские казаки - не включили. Надежность геногеографии: обеспечивается полисистемным подходом - сравнением разных маркеров и данных разных наук.
Ключевые слова: геногеография, Y-хромосома, митохондриальная ДНК, миграции, человечество, Африка, финикийцы, киргизы, русские, поморы, крестоносцы, казаки.
Геногеография - сформировавшаяся наука. Почти столетие назад основатель геногео-графии А.С. Серебровский определил ее как науку историческую, а не биологическую. Он считал, что геногеография, используя генетические маркеры, должна описывать историю формирования популяций и пути миграций человека. Сам А.С. Серебровский в качестве генетического маркера использовал фенотипы кур Дагестана - различия куриных популяций указывали на структуру генофонда их хозяев, на интенсивность обмена генами (и обмена курами) между разными ущельями Дагестана.
За прошедший век изменился инструментарий геногеографии, но ее устремленность к истории стала еще выразительней. В арсенале генетики появились новые мощные маркеры истории популяций - «однородительские» маркеры. Первой завоевала популярность ми-тохондриальная ДНК (мтДНК), передающаяся в поколениях по материнской линии: она позволила убедительно обосновать моноцентристскую теорию происхождения человечества и «выход из Африки» как важнейший этап в расселении по планете человека современного вида. В самый разгар исследований мтДНК, когда на ее
изучении сосредоточились усилия большинства популяционных генетиков, на сцену стремительно вышла другая генетическая система - Y-хро-мосома, наследующаяся в поколениях по отцовской линии. Хотя ей еще не удалось потеснить мтДНК с позиций лидера, Y-хромосома уверенно заняла место рядом с ней. Образовавшийся дуэт стал общепринятым стандартом в мировых исследованиях. В чем же привлекательность этих маркеров? Отсутствие рекомбинации позволяет реконструировать цепочку последовательно происходивших мутаций («от Адама» или же «от Евы»), определить место и время их возникновения, а следовательно - проследить и процесс расселения человечества по планете.
Поэтому современную геногеографию можно назвать наукой об опечатках. Если бы в генетических текстах не было опечаток - мутаций, то геногеографии было бы нечего изучать: все мужчины обладали бы идентичными Y-хромо-сомами, а женщины - идентичными копиями одной и той же молекулы мтДНК. Мутации служат такими же маркерами, как и ошибки переписчиков летописей - благодаря их ошибкам можно дать относительную датировку разных изводов летописей: те изводы, которые включили и старые «опечатки», и свои собственные, рассматриваются как более поздние.
Такой же подход и в геногеографии. По генетическим опечаткам можно построить филогенетическое древо происхождения всех современных генетических линий от одной исходной и выявлять древнейшее генетическое родство населения разных континентов. Наиболее древние мутации будут задавать основные, наиболее крупные ветви древа Y-хромосомы или мтДНК (гаплогруппы). Более поздние мутации показывают, как эти ветви ветвятся на более мелкие (субгаплогруппы). Множество листьев (гаплотипов) различаются только по самым недавним мутациям и одевают все древо, отражая генетическое разнообразие современного человечества.
Если наложить частоты встречаемости различных мутаций на географическую карту, то мы увидим зоны их скопления - те регионы, в которых волей истории эти опечатки размножились. Чем дольше популяция развивалась в этом регионе, тем больше мутаций она могла накопить. Ее дочерние популяции, отправляясь в путь, захватывали с
собой лишь малую часть этого разнообразия. Поэтому мы можем обнаружить и те дочерние регионы, в которые волны миграций занесли те или иные гаплогруппы и гаплотипы. А знание относительного времени возникновения мутаций поможет отделить древние миграции от более поздних.
Конечно, это лишь самые основы, «скелет» того инструмента, которым геногеография отслеживает древние и исторические миграции. Понять его возможности и ограничения легче на живых примерах геногеографических работ. Из огромного массива публикаций мы сделаем случайную выборку - те работы, в которых участвовал наш коллектив. Поскольку они охватывают и целые континенты, и части одного народа, такое беглое рассмотрение позволит охватить большие пространства и времена.
Южная Африка: на заре современного человечества
В популяциях Южной Африки проведен анализ полных нуклеотидных последовательностей митохондриальной ДНК (Behar et al, 2008). Эта трудоемкая работа необходима для ответа на вопрос - каковы были самые первые этапы микроэволюции Homo sapiens. Главным результатом этой работы стало уточнение филогенетического древа человечества (рис. 1). Укажем две важнейшие его черты. Во-первых, разделение древа на два крупных ствола (бушмены и все остальное человечество) произошло по данным о мтДНК более 140 тыс. лет назад. Вторая черта - уже известная из более ранних работ, но от того не менее удивительная. На древе человечества почти все разнообразие приходится на народы Африки. Лишь малая часть ветвей общего древа (показанных на белом фоне) дала побеги на всех остальных континентах - Евразии, Америки, Австралии. Это древо хорошо иллюстрирует общий принцип отслеживания миграций - расселяющиеся популяции, оторвавшиеся от исходного массива, забирают с собой в путь лишь малую часть ветвей, малую часть имеющегося генетического разнообразия. Дальнейшая микроэволюция приводит к росту новых вторичных субгаплогрупп в разных регионах планеты, позволяя прослеживать все более поздние миграции.
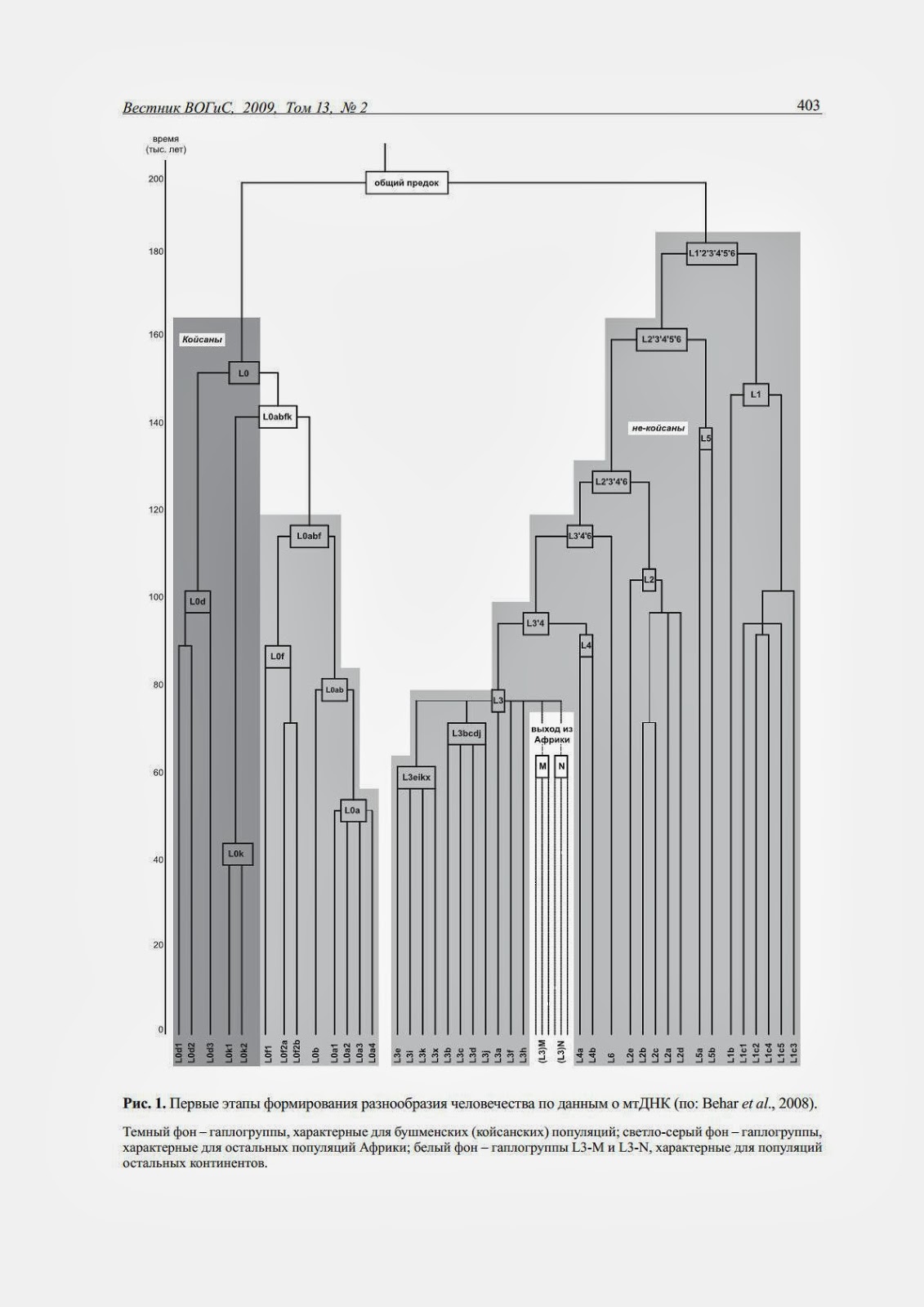
Рис. 1. Первые этапы формирования разнообразия человечества по данным о мтДНК (по: Behar et al., 2008).
Темный фон - гаплогруппы, характерные для бушменских (койсанских) популяций; светло-серый фон - гаплогруппы, характерные для остальных популяций Африки; белый фон - гаплогруппы L3-M и L3-N, характерные для популяций остальных континентов.
Центральная Африка: пигмеи, банту и направление брачных миграций
Следующее важное событие в истории населения Африки и человечества в целом - расхождение генофондов самого низкорослого и самого высокорослого населения планеты - пигмеев и бантуязычных народов. По данным о мтДНК около 70 тыс. лет назад существовала их единая общность (Quintana-Murci et al, 2008). Их разделение, видимо, было связано с климатическими кризисами в истории нашей планеты. Ледниковые периоды в истории земли имели для Африки не менее катастрофичные последствия, чем для Европы. Это было время иссушения планеты -исчезали леса, их место занимали саванны и пустыни, разделившие предков пигмеев и банту. Только много тысяч лет спустя, когда обе популяции приобрели своеобразные антропологические черты, их ареалы вновь стали соприкасаться. Но, как показано, поток генов между ними был строго односторонним: от пигмеев к банту. Народы банту брали в жены маленьких женщин пигмеев, приносивших свои гаплогруппы мтДНК. Но у пигмеев не прослеживаются линии мтДНК бантуязычных народов.
Неолит Европы: палеоДНК и современная изменчивость генофонда
Первая волна заселения Европы связана с палеолитом. Вторая волна - мезолитическая реко-лонизация Европы после отступления ледника. Но больше всего споров вызывает третья волна -неолитических земледельцев. В классическом труде археолога Аммермана и генетика Кавал-ли-Сфорца была сформулирована гипотеза «де-мического распространения»: именно третья -неолитическая - волна расселения земледельцев сформировала основные черты европейского генофонда (Ammerman, Cavalli-Sforza, 1984). Однако впоследствии данные по митохонд-риальной ДНК указали на палеолитический возраст большинства европейских гаплогрупп (Richards et al, 2000), что позволило обосновать гипотезу «культурного распространения»: миграцию земледелия без земледельцев. Оба этих подхода реконструировали генофонды былых эпох по генетической структуре их современных популяций-потомков.
Но лишь данные по древней ДНК (полученные в надежных лабораториях и получившие мировое признание) дают прямую информацию о генофонде древних популяций. Исследование палеоДНК одной из первых в Европе неолитической культуры - линейно-ленточной керамики (Haak et al., 2005) - неожиданно выявило высокую частоту гаплогруппы N1a мтДНК, которая у современных европейцев почти не встречается. Это может означать, что первое земледельческое население Европы действительно почти не оставило потомков. Новые данные, полученные той же группой исследователей в соавторстве с нашим коллективом, позволили уточнить этот вывод: они показали, что эта неолитическая популяция генетически близка к популяциям Передней Азии (Haak et al., unpublished data). Это может означать, что хотя распространение земледелия внутри Европы носило характер «культурной диффузии», появление земледелия в Европе связано с далекой миграцией первых земледельцев, что является своеобразным компромиссом между «демической» и «культурной» гипотезами распространения земледелия.
Финикийцы: расселение по Средиземноморью
Последующие миграции затрагивали уже отдельные регионы Европы. Одной из наиболее ярких миграций являлось расселение первого «морского народа» - финикийцев - из узкой прибрежной полосы по островам и полуостровам в восточной части Средиземноморья. Для того чтобы выявить генетические следы этой миграции, был применен особый прием (Zalloua et al., 2008a). Были изучены такие пары популяций, чтобы одна располагалась на месте исторически документированной финикийской колонии, а другая - в географически близкой местности, предположительно отражая дофи-никийское население. Оказалось, что в каждой паре популяций определенные STR гаплотипы Y-хромосомы указывают на принадлежность к «финикийской» популяции. Детальный анализ геногеографии следов «финикийского гап-лотипа» позволил проследить ход экспансии финикийцев.
Крестоносцы: генетический след в Леванте
Спустя несколько тысячелетий пришло время и для обратной миграции - из Европы на Ближний Восток. Речь идет о крестовых походах. Вопрос о генетических последствиях этих событий оставался открытым - по историческим данным нельзя определить, какая часть населения христианских государств Леванта вернулась в Европу, а какая осталась в Леванте. Но геноге-ография выявила неожиданно четкую картину: у заметной части современного населения Ливана обнаруживается специфический гаплотип Y-хромосомы, который часто встречается у населения западноевропейских стран, но полностью отсутствует в других странах Ближнего Востока (Zalloua et al, 2008b). Так был найден явный генетический отпечаток исторических миграций крестоносцев.
Киргизы: прыжок через Тянь-Шань
Заглянем дальше в Азию - уже не в Переднюю, а в Центральную. Происхождение киргизов - одна из загадок истории. На нынешней территории они появились исторически недавно, а родина их предположительно - енисейские степи. Мы провели экспедиции к двум группам киргизов - Памира и Тянь-Шаня. Оказалось, что при общем сходстве этих генофондов по маркерам Y-хромосомы между ними обнаруживается и примечательное различие: южная - памирская -группа киргизов обладает высокой частотой гаплогруппы N1 с, тогда как у киргизов Тянь-Шаня эта гаплогруппа практически отсутствует (Balanovsky et al., unpublished data). Однако из общего распространения этой «сибирской» гап-логруппы ожидается обратная картина! Анализ STR маркеров показал, что причина именно в миграциях, а не в дрейфе генов. Эти данные указывают на прямые миграции между столь отдаленными регионами, как Южная Сибирь и плоскогорья Восточного Памира.
Русский народ: миграции из Европы и из Азии
Так, гаплогруппа R1a, которая в Европе характерна главным образом для славянских популяций, встречается преимущественно в южных и центральных русских популяциях, а при движении к северу ее частота снижается. И, напротив, при движении в том же направлении от южных к северным русским популяциям постепенно нарастает частота гаплогруппы N1c (N3), которая среди всех европейских популяций чаще всего встречается у народов финской и балтийской групп. Такая картина могла сформироваться в результате постепенной славянской экспансии в северном направлении, сопровождавшейся ассимиляцией автохтонных финно-угорских и балтских племен. В каждом следующем поколении очередной шаг миграции предпринимала смешанная популяция, образовавшаяся в предыдущем поколении, и так постепенно при движении к северу исходный «восточно-славянский» генофонд все в большей мере вбирал в себя автохтонные элементы.
Данные по митохондриальной ДНК, к сожалению, не позволяют нарисовать столь же подробную географическую картину, поскольку степень географической дифференциации для мтДНК значительно меньше, чем для Y-хромо-сомы. Однако рассмотрение митохондриально-го генофонда не отдельных популяций, а всей совокупности данных по русскому населению Центральной России приводит к тем же выводам. Наибольший процент совпадений гаплоти-пов, встреченных в русских популяциях, также характерен и для других восточнославянских народов (украинцев и белорусов) и для разных групп финно-угорских народов (западно-финских и волго-финских). Доля же общих гапло-типов с западными и южными славянами, как и с тюркскими народами Европы, у русских значительно меньше. Это вновь указывает на важную роль финно-угорского субстрата в формировании русского генофонда.
Что же касается второй из крупных миграций, связанных с монгольским завоеванием средневековых русских княжеств, то ее генетические следы обнаружить не удается. И вновь этот вывод взаимно подтверждается анализом и мтДНК, и Y-хромосомы, и данными антропологии. Например, суммарная частота восточно-евразийских гаплогрупп мтДНК в русских популяциях не достигает и 2 %: эта же частота характерна и для западноевропейских народов. Для Y-хромосомы типичным «монгольским» маркером является гаплогруппа С (ее носителем был, как считается, Чингисхан, - эта гаплогруппа является самой частой у монголов и родственных им народов). Однако в русских популяциях гаплогруппа С практически не встречена (частота не достигает 1 %, т. е. с формально-генетических позиций полиморфизм по этому признаку в русских популяциях отсутствует, и русское население может считаться полностью «генетически европейским»).
Северные русские: пенеги
| |||

Рис. 2. Структура русского генофонда по маркерам Y-хромосомы (по: Balanovsky et al., 2008).
На рис. 2 показано взаимное генетическое сходство по маркерам Y-хромосомы русских популяций из различных частей исторического реала. Легко заметить, что все южные и центральные русские популяции генетически похожи и формируют единый гомогенный кластер, тогда как разные популяции Русского Севера значительно отличаются как от южноцентрального кластера, так и друг от друга. Такие большие различия между северными популяциями являются хорошей иллюстрацией действия дрейфа генов. Спектр митохондри-альных гаплогрупп в северных популяциях в основном воспроизводит усредненный об
щерусский спектр, свидетельствуя об общем корне всех: южных, центральных и северных русских популяций. Можно полагать, что эти гаплогруппы были принесены на север первыми русскими поселенцами в 12-13 вв. и до сих пор сохраняются у их потомков. В то же время на севере с небольшими частотами встречены и нетипичные для русских гаплогруппы A и D5, которые встречаются у соседних финно-угорских народов северо-востока Европы. Очевидно, что здесь мы снова видим генетическое свидетельство ассимиляции мигрировавшими сюда славянами автохтонного финно-угорского населения. Но мы видим больше: не только схему формирования этой северно-русской популяции, но и последующую историю ее генофонда, протекавшую из-за малого пресса миграций под знаком генетического дрейфа. И видим мы это опять-таки по спектру гаплогрупп, который на Русском Севере, оказывается, хотя и сходен с общерусским, но значительно обеднен: многие гаплогруппы, представленные в Центральной России, на Русском Севере исчезли в результате случайного дрейфа.
Генофонды различных групп казачества: контрасты
Совершенно иначе складывалась судьба русского генофонда на противоположной - южной -окраине русского ареала. Казачьи группы, сформировавшиеся на границах со степными и горскими народами Предкавказья и Северного Кавказа, не испытывали значительного дрейфа генов. Зато они в полной мере подвергались влиянию миграций в ходе богатой событиями истории этого региона. Наиболее любопытные результаты получены при анализе полиморфизма Y-хромосомы у терских и кубанских казаков (Balanovsky et al, unpublished data). Терское казачество является самым первым на Кавказе и исходно было немногочисленным. Напротив, кубанское казачество сформировалось исторически поздно (в 19 в.) за счет массовых миграций с Дона, а также внутренних областей России и Украины. К этим историческим сведениям генетическое исследование добавило новую любопытную черту. Оказалось, что терское казачество вобрало в себя более четверти местных, автохтонных кавказских гаплотипов Y- хромосомы. Кубанское же казачество не имеет генетического сходства с кавказскими народами (хотя проживает с ними бок о бок уже два века), и его генетический портрет совпадает с портретом населения Южной России и Украины. Можно полагать, что в ходе раннего периода жизни терских казаков на Кавказе их отношения с окружающими народами были более мирными, чем впоследствии в период Кавказской войны, и терские казаки включили большое число мужчин - выходцев из окружающих горских народов.
Надежность генетики -сравнение итогов разных наук
Даже такой беглый обзор разворачивает внушительную панораму миграций популяций человека, в изучении которых большую роль сыграли данные популяционной генетики: от древнего отделения бушменской ветви человечества 140 тыс. лет назад в Южной Африке до формирования кубанских казаков полтора века назад на Северном Кавказе. Такие успехи генетики в изучении доисторических, исторических и современных миграций обеспечили ей большой авторитет в глазах смежных, особенно гуманитарных наук, таких, как археология, сравнительная лингвистика и отчасти физическая антропология.
Но большой авторитет накладывает на нас и большую ответственность за достоверность и обоснованность наших выводов. К сожалению, не всегда генетикам удается устоять перед соблазном поспешных интерпретаций и ярких выводов. И наиболее трезво мыслящие гуманитарии замечают ту легкость, с которой выводы генетиков меняются на противоположные всего лишь через несколько лет. Что же делать, как отличить достоверные выводы генетических исследований (которые уже не изменятся при появлении новых данных или новых методов их анализа) от выводов сиюминутных и шатких? Это задача непростая, но крайне важная.
Один из возможных ответов состоит в тщательном следовании хорошо известным правилам популяционно-генетического обследования: достаточного объема выборок (более 70 человек для аутосомных и более 150 человек для однородительских - гаплоидных - марке
ров); репрезентативности выборок (включение только неродственных индивидов, относящихся к коренному населению популяции вплоть до третьего поколения) и другим.
Второй, дополняющий первый, путь к повышению надежности наших выводов - во взаимном подтверждении результатов, полученных при использовании разных маркеров.
Важным шагом на этом пути является одновременное использование данных по мтДНК и Y- хромосоме: при этом надежными следует признавать только те результаты, которые подтверждены обеими системами. Однако обе эти системы в сущности очень похожи: обе гаплоидны, не рекомбинируют, анализируются одними и теми же филогеографическими методами, наиболее уязвимы для действия дрейфа генов. А это может привести к искажениям реконструируемой картины миграций. Поэтому следующий шаг - это расширение спектра анализируемых генетических систем за счет аутосомных ДНК и классических генных маркеров, а также включения информативных квазигенетических систем - фамилий, антропологических, археологических и лингвистических признаков. Когда картины мира - русского, европейского, евразийского -совпадают вопреки тому, что они обрисованы совершенно разными свидетелями (генетикой, антропонимикой, антропологией), мы можем быть уверены, что генетические следы миграций реальны и достоверны.
Использование многих систем - полисистемный подход - не только позволяет достичь нового уровня надежности результатов генетических исследований и тем самым укрепить авторитет генетики в глазах смежных наук, но и открывает путь к реальному синтезу знаний об истории популяций человека, полученных самыми разными науками.
Литература
Балановская Е.В., Балановский О.П. Русский генофонд на Русской равнине. М.: Луч, 2007а. 416 с.
Балановская Е.В., Балановский О.П. Русский генофонд: свидетельства «очевидцев» // Наука в России. 2007б. № 2. С. 54-63.
Ammerman A.J., Cavalli-Sforza L.L. Neolithic Transition and the Genetics of Populations in Europe. Princeton. N.J.: Princeton Univ. Press, 1984.
Balanovsky O., Rootsi S., Pshenichnov A. et al. Two sources of the Russian patrilineal heritage in their Eurasian
context // Amer. J. Hum. Genet. 2008. V 82. № 1. P. 236-250.
Behar D.M., Villems R., Soodyall H. et al. The dawn of human matrilineal diversity // Amer. J. Hum. Genet. 2008. V. 82. № 5. P. 1130-1140.
Haak W., Forster P., Bramanti B. et al. Ancient DNA from the first European farmers in 7500-year-old
Neolithic sites // Science. 2005. V. 310. № 5750.
P. 1016-1018.
Quintana-Murci L., Quach H., Harmant C. et al. Maternal traces of deep common ancestry and asymmetric gene flow between Pygmy hunter-gatherers and Bantu-speaking farmers // Proc. Natl Acad. Sci. USA. 2008. V. 105. № 5. P. 1596-1601.
Richards M., Macaulay V., Hickey E. et al. Tracing European founder lineages in the Near Eastern
mtDNA pool // Amer. J. Hum. Genet. 2000. V. 67.
№ 5. P. 1251-1276. Zalloua P.A., Platt D.E., El Sibai M. et al. Identifying genetic traces of historical expansions: Phoenician footprints in the Mediterranean // Amer. J. Hum.
Genet. 2008a. V. 83. I. 5. P. 633-642.
Zalloua P.A., Xue Y., Khalife J. et al. Y-chromosomal diversity in Lebanon is structured by recent historical
events // Amer. J. Hum. Genet. 2008b. V. 82. № 4. P. 873-882.