«Как определить кто русский? Вот генетики сейчас проводят исследования и вроде у них получается, что самые чистокровные славяне – это поляки, а в русских много тюркской крови...»
Какой бы признак мы ни взяли, видим: русские – типичные европейцы, а азиатское завоевание оставило след в их истории, но не в генофонде. Не беремся говорить о России и русских в культурном, историческом, гуманитарном смысле. Однако биологически их генофонд не является промежуточным между европейским и азиатским, он – типичный европейский. Уточним: самый восточный из этого ряда, стоящий «на передовой». Отдельные азиатские влияния прослеживаются в нём больше, чем в западных соседях. Но базовый, главный вывод, который следует из проведенного исследования – практически полное отсутствие в русском генофонде монголоидного вклада
Елена БАЛАНОВСКАЯ, Олег БАЛАНОВСКИЙ
Доля восточно-евразийских гаплогрупп в русском населении составила лишь 2%. То есть очень малую величину. Почти столь же малую, что и в генофонде поляков (1,5%) или севера Европы, где уже точно никаких «татаро-монгольских вкладов» не было (у норвежцев 0,6%, у исландцев 0,7%, у карел 4,8% и т.д.). Средняя «фоновая» частота восточно-евразийских гаплогрупп в Европе равна 3,6%. То есть в русском генофонде (2,0%) она даже меньше, чем «средняя по Европе», поэтому монголоидный компонент у русских оказывается не просто нулевым, но даже с отрицательным знаком. Итак, мы не видим последствий монгольского нашествия в русском генофонде – или тогда должны видеть эти последствия и в польском, и в норвежском, и в карельском, и в других генофондах Европы
http://dienekes.blogspot.com/2008/04/origins-of-mongoloid-mtdna-in-slavs.html
См также дополнительная информация.
http://www.gazeta.ru/science/2008/01/14_a_2552231.shtml
 Теперь доступно и подробно по делению всего исходного материала на кластеры количеством от 3 до 11. Структурная диаграмма, построенная на основе SNP-маркёров, представляет собой массив исходных данных, где каждый столбец составляет отдельный образец, либо — в данном случае — сгруппированные в популяции средние значения по образцам. Деление на кластеры проводится машинным анализом сходства фрагментов SNP-маркёров при заранее заданном количестве кластеров. Другими словами, схожие последовательности отдельных образцов объединяются в кластеры и обозначаются своими цветами.
Теперь доступно и подробно по делению всего исходного материала на кластеры количеством от 3 до 11. Структурная диаграмма, построенная на основе SNP-маркёров, представляет собой массив исходных данных, где каждый столбец составляет отдельный образец, либо — в данном случае — сгруппированные в популяции средние значения по образцам. Деление на кластеры проводится машинным анализом сходства фрагментов SNP-маркёров при заранее заданном количестве кластеров. Другими словами, схожие последовательности отдельных образцов объединяются в кластеры и обозначаются своими цветами.
 http://dienekes.blogspot.ru/2011/04/genetic-structure-of-west-eurasians.html
http://dienekes.blogspot.ru/2011/04/genetic-structure-of-west-eurasians.html
Ну и где шведы - нордманы, якобы основатели русского государства ??????? Вот синтетическая карта Европы, показывающая распределение и родство различных этнических групп:


Популяционная генетика человека включает: 1) Y - хромосома (см. Вики) - мужская линия.

2) Х - хромосома (см. Вики) - женская линия. 3) Аутосомы (см.Вики) - не половые хромосомы.


EUROPEAN GENETIC VARIATION - Аутосомные маркеры о русских
 THE FIRST component is almost superimposable to the archaeological dates of the spread of farming from the Middle East between 10,000 and 6,000 years ago.
THE FIRST component is almost superimposable to the archaeological dates of the spread of farming from the Middle East between 10,000 and 6,000 years ago.
 THE SECOND principal component parallels a probable spread of Uralic people and/or languages to the northeast of Europe [c. 4,000 years ago].
THE SECOND principal component parallels a probable spread of Uralic people and/or languages to the northeast of Europe [c. 4,000 years ago].
 THE THIRD is very similar to the spread of pastoral nomads (and their successors) who domesticated the horse in the steppe towards the end of the farming expansion, and are believed by some archaeologists and linguists to have spread most Indo-European languages to Europe.
THE THIRD is very similar to the spread of pastoral nomads (and their successors) who domesticated the horse in the steppe towards the end of the farming expansion, and are believed by some archaeologists and linguists to have spread most Indo-European languages to Europe.
 THE FOURTH is strongly reminiscent of Greek colonization in the first millennium B.C.
THE FOURTH is strongly reminiscent of Greek colonization in the first millennium B.C.
 THE FIFTH corresponds to the progressive retreat of the boundary of the Basque language. Basques have retained, in addition to their language, believed to be descended from an original language spoken in Europe, some of their original genetic characteristics.
THE FIFTH corresponds to the progressive retreat of the boundary of the Basque language. Basques have retained, in addition to their language, believed to be descended from an original language spoken in Europe, some of their original genetic characteristics.
Лужичане, Поляки, Украинцы, Белорусы, Венгры, Чехи, Словаки, Латыши, Литовцы, Киргизы, Алтайцы, Таджики, Ишкашим, Белуджи, Пенджабцы, Пуштуны. Однотипные Хромосомы сгруппированы в гаплоидные группы, - свидельствующие о родстве народов, всего их 18. Вот например, некоторые из них.



http://www.eupedia.com/europe/european_y-dna_haplogroups.shtml
Общепринятые Cубклады гаплогруппы R1a


Изменчивость на разных уровнях популяционной иерархии.
Карта западно-евразийской гаплогруппы R1a показывает ее доминирование в
Восточной Европе и высокие частоты также в горах Алтая и Тянь-Шаня (где снижены
частоты восточно-евразийских гаплогрупп). Такая «альтернативность» гаплогруппы R1a и
восточно-евразийских гаплогрупп создает необычный кольцевой ареал R1a.
Филогенетическая сеть изменчивости этой гаплогруппы (рис. ниже ) подтверждает
существование древних кластеров STR-гаплотипов R1a в южносибирских популяциях.
Можно предполагать, что первоначально R1a была распространена по степной полосе
вплоть до Монголии. Эта зона согласуется с ареалом скифо-сарматских групп.
 Что подтверждается и картой, где выявлены очаги носителей гаплоидной группы R1a1а
Что подтверждается и картой, где выявлены очаги носителей гаплоидной группы R1a1а
 См.http://en.wikipedia.org/wiki/Proto-Indo-Europeans
См.http://en.wikipedia.org/wiki/Proto-Indo-Europeans
![I_Distribution[1]](http://ic.pics.livejournal.com/rugerik/28795279/95957/95957_original.jpg)
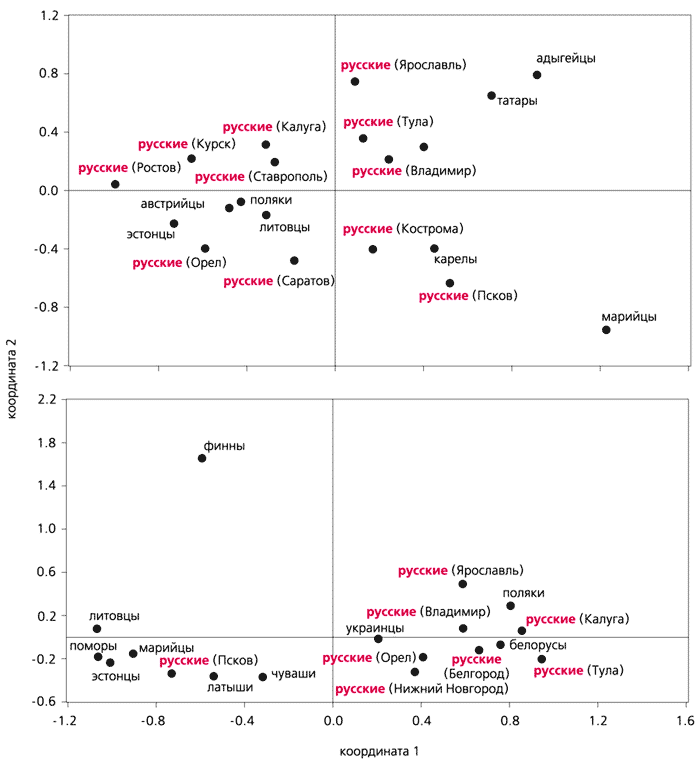
Обнаружение четких географических закономерностей, с одной стороны, развивает тенденции, показанные ранее . С другой стороны, основной вывод этих авторов состоял в том, что географическое соседство, а не лингвистическое родство задает структуру генофонда. Результаты исследования, О.Балановского подтверждая важность
географического фактора, указывают, что доминирует структурированность по
лингвистическому принципу. Это следует из результатов многомерного шкалирования. См.Рис.

В этот вид анализа включены те народы, для которых имелись данные по четырем или более региональным популяциям с объемом выборки не менее 70 человек.
В данном случае этнический фактор совпадает с лингвистическим, поскольку этническая принадлежность популяции определялась по языку. График показывает, что во всех случаях популяции, относящиеся к одному народу, кластеризуются совместно, образуя «этнические облака», почти не перекрывающиеся друг с другом. См.рис выше.
Два исключения касаются славянских групп. Во-первых, северные русские
популяции не присоединяются к южным, а образуют отдельный кластер. Во-вторых,
популяции поляков, южных русских и украинцев формируют не три отдельных, а одно единое «этническое облако», демонстрируя общность генофонда. Таким образом, славяноязычные популяции, занимающие половину территории Европы, оказываются генетически не различными (как должно бы быть в случае доминирования в структуре генофонда географического фактора), а сходными, в согласии с лингвистическими данными о близком родстве восточнославянских и западнославянских языков

Поскольку этнический фактор оказался столь существенным для структурирования
европейского генофонда, был проведен анализ межпопуляционной изменчивости на двух уровнях популяционной иерархии: а) внутриэтническом (различия популяций в пределах одного народа, т.е. оценка гетерогенности этнического генофонда); б) межэтническом (различия между усредненными этническими частотами гаплогрупп разных народов Европы).

Таблица показывает, что в пределах Европы наибольшая гетерогенность
характерна для финнов, хорватов, русских и итальянцев; шведы и немцы демонстрируют умеренную гетерогенность; другие народы (греки, турки, поляки, белорусы, украинцы)
характеризуются сниженной в масштабе Европы гетерогенностью. В среднем,
гетерогенность типичного европейского народа составляет 0.03 (Gst=0.029, Fst=0.027).
Таким образом, по гаплогруппам Y-хромосомы для населения Европы различия между популяциями одного этноса значительно меньше, чем различия этносов между собой (межэтнический уровень Gst =0.15, Fst =0.17). Общая величина межпопуляционных различий
по Y-хромосоме (суммарно внутриэтнический и межэтнический уровни) составляет для Европы около 0.19. Это является дополнительным подтверждением резко выраженных
различий внутри Европы по Y-хромосоме , поскольку по другим генетическим системам межпопуляционная изменчивость Европы на порядок ниже: Gst =0.02.
Балановский О.П. ИЗМЕНЧИВОСТЬ ГЕНОФОНДА В ПРОСТРАНСТВЕ И ВРЕМЕНИ:СИНТЕЗ ДАННЫХ О ГЕНОГЕОГРАФИИ МИТОХОНДРИАЛЬНОЙ ДНК И Y-ХРОМОСОМЫ.
Родственники русских по Доминируюшей икс хромосоме мтДНК.


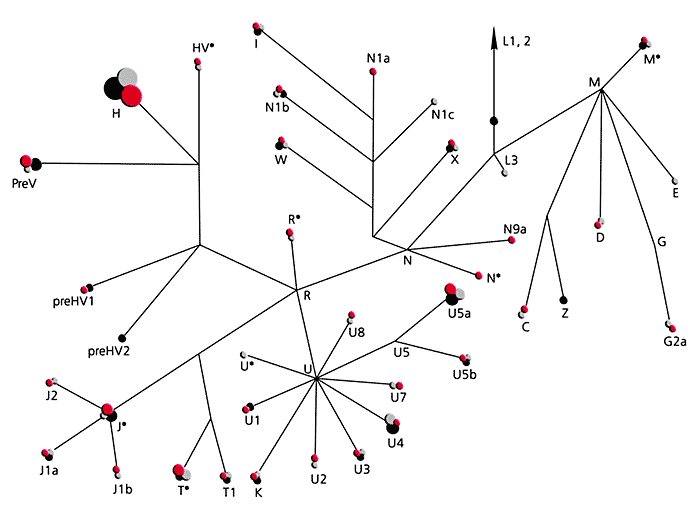
При сравнительном изучении мтДНК у народов выявленные гаплотипы были отнесены к определенным линиям в соответствии с классификацией Макуолли. Для большинства расовых и географических групп установлена специфичность типов мтДНК (рис. ниже). Более 90% всех типов мтДНК народов Западной Евразии принадлежит к линиям Н, J, V, 1, К, Т, U, W и X, которые рассматриваются как потомки европейского генного пула, существовавшего в верхнем палеолите. Митохондриальный геном азиатских популяций включает два суперкластера М и N, которые соединяются в африканском макрокластере L3. Последний рассматривается в качестве общего предка для всех неафриканских популяций мира. Около 50% линий мтДНК коренных жителей Азии принадлежит к суперкластеру М, который подразделяется на линии С, Z, D, G, Е и др.
. Суперкластер N в Азии делится на несколько линий: А, Y, В, F




Следы смешения народов можно найти в их генах. На рисунке частоты европейских и азиатских линий мтДНК для некоторых народов, населяющих Россию.
 Идентификация митоДНК у русских, европейская она или азиатская.
Идентификация митоДНК у русских, европейская она или азиатская.
 Женские гаплогруппы у русских также являются полностью славянскими, о чем свидетельствует их сравнение с теми же гаплогруппами у поляков.
Женские гаплогруппы у русских также являются полностью славянскими, о чем свидетельствует их сравнение с теми же гаплогруппами у поляков.

 Cм. http://aquilaaquilonis.livejournal.com/18058.html
Cм. http://aquilaaquilonis.livejournal.com/18058.html
Подобное же единообразие демонстрирует и сравнение МитоДНК у русского и немецкого народов. Данные взяты из Европедии. Явно генетика у немок славянская, что позволяет сделать некоторые выводы .....

ИЗМЕНЧИВОСТЬ МИТОХОНДРИАЛЬНОЙ ДНК В ЕВРОПЕ
Изменчивость мтДНК изучена в 22 популяциях русских, украинцев и белорусов,
охватывающих все части обширного ареала восточных славян.

Частоты гаплогрупп приведены в таблице ниже. Данные представлены также на сайте www.genofond.ru .Использование этой базы данных MURKA расширяет число доступных для анализа популяций (см. табл. ).

Такое обилие данных позволило провести строгий отбор популяций по следующим критериям: а) объем выборки не менее 70 (меньшие выборки были исключены или объединены); б) надежное определение гаплогрупп (подтвержденных ПДРФ-анализом или однозначно прогнозируемых по ГВС1); в) наличие четкой географической привязки в описании выборки. Был также проведен и строгий отбор гаплогрупп: в анализ включены только гаплогруппы, средняя частота которых в Европе не менее 1% (критерий полиморфизма).

Перечисленным критериям удовлетворяли 118 популяций (средний объем выборки N=200, суммарно 23500 образцов) и 35 гаплогрупп.
КЛАСТЕРИЗАЦИЯ ПОПУЛЯЦИЙ ПО ЧАСТОТАМ ГАПЛОГРУПП.
При анализе методом многомерного шкалирования (рис. А) четко выделились четыре кластера. Первый кластер включил только саамов, что не удивительно, учитывая их генетическое своеобразие. Второй кластер включил те популяции восточных рубежей Европы, у которых повышена частота восточно-евразийских гаплогрупп.
Третий кластер включил популяции Передней Азии и Кавказа. Все остальные популяции с
основной территории Европы (от Волги до Пиренейского полуострова) вошли в четвертый
«паневропейский» кластер, небольшие размеры которого на графике свидетельствуют о низкой межпопуляционной изменчивости. Эти результаты подтверждают гомогенность генофонда Европы , но указывают на своеобразие генофондов Приуралья и Передней Азии.
При отдельном анализе только тех популяций, которые вошли в «паневропейский»
кластер (рис. Б), обнаружилась обособленность популяций северо-востока Европы (главным образом, финно-угорских). Однако подавляющее большинство популяций по-прежнему группировалось в единый гомогенный кластер, который можно назвать «узко-европейским» (рис. Б).
 Наличие обширной базы данных позволило провести отдельный анализ и этого кластера. В результате (рис. В) такого «рассмотрения под большим увеличением» в пределах «узко-
Наличие обширной базы данных позволило провести отдельный анализ и этого кластера. В результате (рис. В) такого «рассмотрения под большим увеличением» в пределах «узко-
европейского» кластера выявился целый ряд кластеров второго порядка. Популяции,
говорящие на языках одной лингвистической группы, расположились в генетическом
пространстве рядом друг с другом. Так, популяции германской, славянской, романской и балтской языковых групп формируют каждая отдельную группу и в генетическом
пространстве. Учитывая, что население Шотландии лишь исторически недавно сменило язык, их генетическое сходство с ирландцами обрисовывает общность митохондриального генофонда и кельтской группы (рис. В).
В обнаруженной закономерности генофонда Европы (популяции одной языковой
группы характеризуются сходными митохондриальными генофондами) отмечаются два исключения: румыны и аромуны (лингвистически принадлежат к романской группе, но генетически обособлены), а также эстонцы и венгры (лингвистически финно-угорская группа, но генетически примыкают к «славянскому» кластеру).
РОЛЬ ЛИНГВИСТИЧЕСКОГО И ГЕОГРАФИЧЕСКОГО ФАКТОРОВ.
Значимость географического и лингвистического факторов оценивалась во многих работах . Впервые обнаруженная значимость лингвистической принадлежности популяций для структурирования митохондриального генофонда Европы подтверждена и другим методом: проведен анализ связи генетических, географических и лингвистических расстояний между популяциями Европы (тест Мантеля, табл. ниже). Выявлен примерно одинаковый уровень корреляции генетических расстояний и с географическими, и с лингвистическими расстояниями.

ИЗМЕНЧИВОСТЬ В ПРЕДЕЛАХ ЛИНГВИСТИЧЕСКИХ ГРУПП.
Для уточнения выявленной связи генетических кластеров популяций с лингвистическими группами была рассчитана межпопуляционная изменчивость (гетерогенность) в пределах каждой из лингвистических групп Европы (табл. , рис. ).
 Оказалось, что митохондриальные генофонды всех лингвистических групп индоевропейской семьи характеризуются умеренной гетерогенностью (от 0.01 до 0.04), тогда как гетерогенность финно-угорской группы уральской языковой семьи значительно выше (0.42 до коррекции, 0.07 после исключения саамов и удмуртов). Это согласуется с обособленностью генофонда финно-угорских популяций и их дисперсным расположением на графике многомерного шкалирования (рис. Б). Значительная гетерогенность тюркских народов Европы (0.24) может быть вызвана предполагаемой для многих из них (турки, чуваши и др.) сменой автохтонного языка на тюркский без значительных изменений генофонда.
Оказалось, что митохондриальные генофонды всех лингвистических групп индоевропейской семьи характеризуются умеренной гетерогенностью (от 0.01 до 0.04), тогда как гетерогенность финно-угорской группы уральской языковой семьи значительно выше (0.42 до коррекции, 0.07 после исключения саамов и удмуртов). Это согласуется с обособленностью генофонда финно-угорских популяций и их дисперсным расположением на графике многомерного шкалирования (рис. Б). Значительная гетерогенность тюркских народов Европы (0.24) может быть вызвана предполагаемой для многих из них (турки, чуваши и др.) сменой автохтонного языка на тюркский без значительных изменений генофонда.
ГАПЛОТИПИЧЕСКОЕ РАЗНООБРАЗИЕ мтДНК.
Все выше описанные виды анализа
проведены по частотам гаплогрупп. Для увеличения разрешающей способности проведен также анализ и по отдельным гаплотипам. Уровень гаплотипического разнообразия рассчитан для 51 восточноевропейских популяций как по собственным данным, так и по созданной базе данных . Столь представительный анализ гаплотипического разнообразия мтДНК выполнен впервые в мире. Картографирование этого показателя демонстрирует закономерное убывание к северу: от 0.99 у популяций Причерноморья до 0.95 и ниже на берегах Северного Ледовитого океана (рис. ниже).
 Снижение внутрипопуляционного разнообразия является индикатором действия дрейфа генов, особенно сильного в небольших по размеру и изолированных популяциях севера Европы. Эту зону (см. рис. выше) населяют в основном финно-угорские народы, межпопуляционное разнообразие которых велико. Итак, низкое внутрипопуляционное и высокое межпопуляционное разнообразие оказываются двумя сторонами одной медали, обусловленными действием дрейфа генов (эффект Валунда) в подразделенном финно-угорском генофонде. Северные области способны поддерживать меньшую численность населения, к тому же северные популяции более изолированы за счет меньшей проходимости территории. Такая эколого-географическая трактовка подтверждается корреляцией карты разнообразия (см. рис. выше) с картой плотности населения, которая также снижается к северу.
Снижение внутрипопуляционного разнообразия является индикатором действия дрейфа генов, особенно сильного в небольших по размеру и изолированных популяциях севера Европы. Эту зону (см. рис. выше) населяют в основном финно-угорские народы, межпопуляционное разнообразие которых велико. Итак, низкое внутрипопуляционное и высокое межпопуляционное разнообразие оказываются двумя сторонами одной медали, обусловленными действием дрейфа генов (эффект Валунда) в подразделенном финно-угорском генофонде. Северные области способны поддерживать меньшую численность населения, к тому же северные популяции более изолированы за счет меньшей проходимости территории. Такая эколого-географическая трактовка подтверждается корреляцией карты разнообразия (см. рис. выше) с картой плотности населения, которая также снижается к северу.
Можно предположить причинно-следственную цепь взаимосвязанных изменений от юга к северу сразу нескольких параметров: географической широты (возрастает),
продуктивности среды (убывает), плотности населения (убывает), эффективного размера
популяции (убывает), изоляции субпопуляций (возрастает), дрейфа генов (возрастает),
гаплотипического разнообразия мтДНК (убывает).
Согласно теории популяционной генетики интенсивный дрейф генов должен приводить
к возрастанию случайного инбридинга, а, следовательно, и генетического груза популяции. И действительно, выявлена достоверная корреляция (r=0.4) между гаплотипическим разнообразием мтДНК и отягощенностью популяций аутосомно-рецессивной наследственной патологией . Это подтверждает роль дрейфа генов как ключевого фактора в формировании северных генофондов.
ОСОБЕННОСТИ ГЕНОФОНДА ВОСТОЧНЫХ СЛАВЯН
Структура генофонда изучена как на уровне Европы и Евразии в целом, так и на более
низких иерархических уровнях. В качестве такого модельного объекта выбраны восточные
славяне (наиболее многочисленная группа населения России и сопредельных стран
Европы), а среди восточных славян особое внимание уделено русскому этносу.
МИТОХОНДРИАЛЬНЫЙ ГЕНОФОНД ВОСТОЧНЫХ СЛАВЯН.
Исследованию изменчивости мтДНК у восточных славян в их историческом ареале посвящен ряд работ , различавшихся подходами к формированию выборок. Поскольку для близкородственных генофондов этот вопрос особенно важен, то в анализ включены главным образом собственные данные О.Балановского. Результаты анализа частот гаплогрупп у 22 восточнославянских популяций (N=2 396 образцов) и окружающих их неславянских народов представлены на рис. ниже.

Во избежание перегрузки графика, на нем оставлены лишь популяции (от Балканского полуострова до Урала), имеющие непосредственное отношение к этногенезу восточной ветви славян. Выявленная структура генофонда сводится к трем кластерам. В «приуральский» кластер вошли восточные финно-угорские (марийцы, коми) и тюркские народы (татары, чуваши). В «восточно-европейский» кластер вошли западные и восточные славяне, балты, а также балканские народы. В «северо-европейский» кластер вошли западные финно-угорские народы и северные русские популяции.
Таким образом, выявляется генетически гомогенная балто-славянская общность, к
которой тяготеют также и балканские популяции («восточно-европейский» кластер). От этой общности отличаются финно-угорские и тюркские народы Восточной Европы, генофонд которых, в свою очередь, подразделяется на западную и восточную части (кластеры 1 и 3).
Такая картина могла сформироваться в ходе славянской колонизации Восточно-Европейской равнины : двигающееся с запада славянское население несло с собой «восточно-европейский» митохондриальный генофонд, который замещал ранее существовавший на этих территориях автохтонный «приуральский» генофонд, разделяя оставшиеся не ассимилированными группы на западную и восточную части.
ГЕНОФОНД ВОСТОЧНЫХ СЛАВЯН ПО Y-ХРОМОСОМЕ.
Анализ Y-хромосомы обрисовал в целом аналогичную картину: восточнославянские популяции оказываются сходными между собой и с западнославянскими популяциями, при этом прослеживается своеобразие северных русских популяций . Различия в результатах, полученных по мтДНК и Y-хромосоме, невелики и связаны
главным образом с тем, что мтДНК в целом более гомогенна в Европе и поэтому в меньшей
степени, чем Y-хромосома, выявляет различия между генофондами. Например, по мтДНК
южные славяне присоединяются к остальной славянской общности, хотя в генетическом
пространстве располагаются на ее окраине (рис. выше). Но результаты по Y-хромосоме
(разрешающая способность которой выше) уточняют картину: четко видно, что южные
славяне отделены от восточных и западных славян . Ряд закономерностей выявлен для генофондов украинцев и белорусов. Литературные данные об этих популяциях получены по трудно сопоставимым панелям SNP маркеров, поэтому мы опирались главным образом на собственные данные. Для украинцев показано сходство с более северными группами (Польши, Белоруссии и юга России) и некоторое своеобразие западных украинцев. Обнаружена гомогенность генофонда белорусов, сквозь которую все же пробиваются слабые различия между северными и южными группами .
СТРУКТУРА РУССКОГО ГЕНОФОНДА.
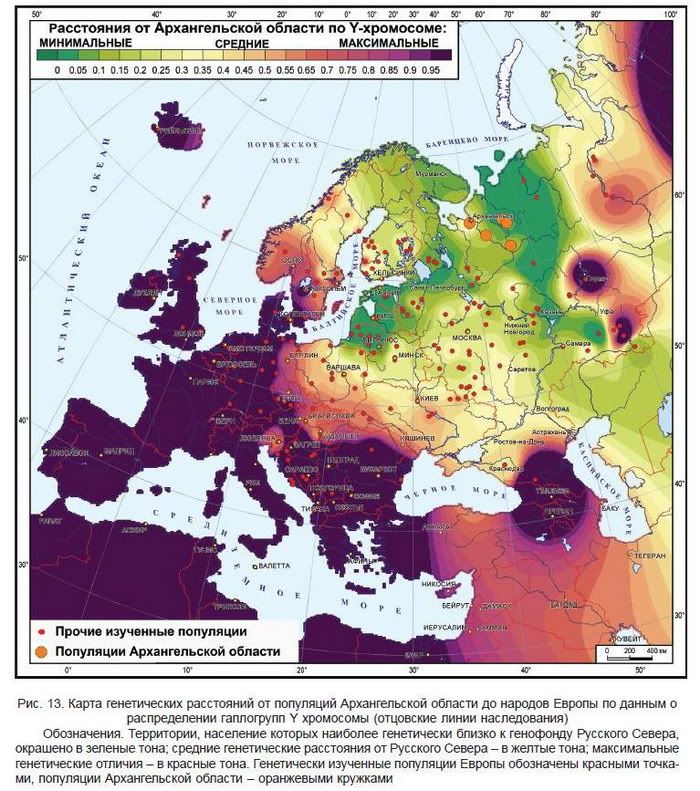
Более высокая разрешающая способность Y-хромосомы по сравнению с мтДНК обнаруживается и при анализе русского генофонда. По мтДНК его структура выявляется лишь на уровне тенденций: в изменении частот некоторых гаплогрупп с юга на север, в отличии окраинных популяций казаков и поморов (Балановский и др., 2009). Близкие результаты получены и другими авторами . При анализе же Y-хромосомы изменчивость большинства гаплогрупп близка к клинальной, что подтверждено анализом автокорреляций . Информативность Y-хромосомы проявляется и при многомерном анализе: центральные и южные популяции оказываются генетически сходными, а северные популяции - своеобразными (рис. ниже).

Кардинальные различия северных и южных русских проявляются и при анализе карт
главных компонент изменчивости различных признаков: гаплогрупп Y-хромосомы и
классических маркеров , фамилий ,
антропологических признаков . Взаимное подтверждение результатов по четырем системам признаков (принцип «полисистемного подхода») не
оставляет сомнений в реальности обнаруженной закономерности широтной изменчивости русского генофонда.
Первая причина высокой межпопуляционной изменчивости на Русском Севере - сниженная плотность населения и меньший эффективный размер популяций . Поэтому дрейф генов в
северных русских популяциях действует сильнее. Тем самым снова обнаруживается
параллелизм с изменчивостью мтДНК (с описанным выше широтным трендом
гаплотипического разнообразия и связанных с ним параметров). Но отличия северных
русских популяций вызваны не только действием дрейфа генов: в этом случае северные русские группы разместились бы случайным образом вокруг южных, без систематического сдвига в одну сторону. Поэтому второй причиной могут быть особенности происхождения северных русских. Это побудило нас провести специальное исследование генофонда Русского Севера .
ПРОИСХОЖДЕНИЕ СЕВЕРНЫХ РУССКИХ ПОПУЛЯЦИЙ.

Анализ популяций Русского Севера проведен по совокупности данных о полиморфизме Y-хромосомы, мтДНК, аутосомных маркеров и фамилий. По данным о Y-хромосоме северные русские вошли в обширный «северный» кластер вместе с популяциями балтов (латышей и литовцев), финноязычных народов (коми, финнов, эстонцев, карелов), а также германоязычных шведов.
Диаграмма-график родства русского народа со скандинавами и конкретно со шведами.

http://www.ncbi.nlm.nih.gov/pmc/articles/PMC2730349/?tool=pubmed
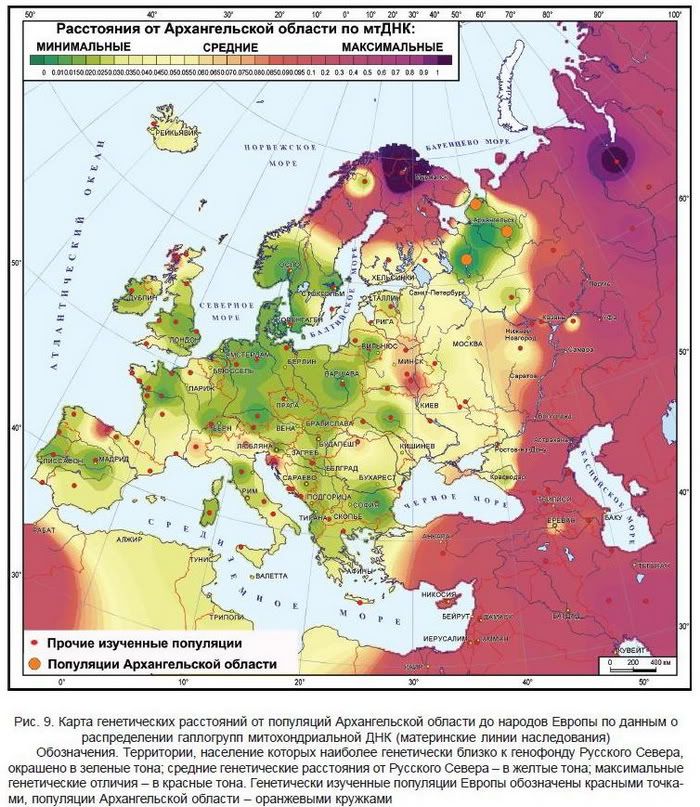
Сходство Русского Севера с географически отдаленными балтами более выражено, чем с финно-уграми. Этот пестрый в лингвистическом отношении (славяне, балты, финно-угры, германцы), но географически единый (от Балтики до Печоры) континуум популяций мог сформироваться еще до выделения балто-славянских и финских лингвистических групп. По митохондриальной ДНК Русский Север обнаруживает близость с еще более отдаленными популяциями Европы (рис. ниже).

Русский Север оказался генетически близким к
норвежцам, немцам, полякам, литовцам и т.д., и при этом генетически отличным от
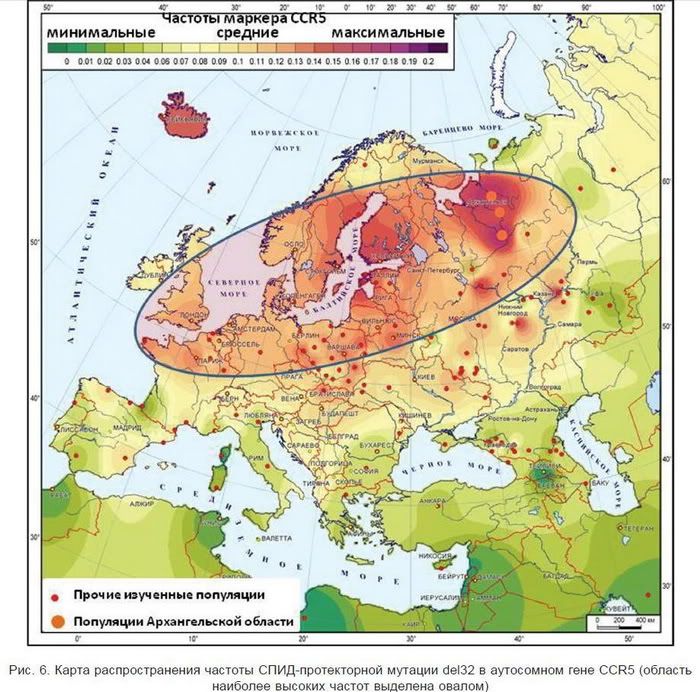
финноязычных популяций. Анализ аутосомного маркера CCR5del32, изменчивость которого активно изучалась многими российскими коллективами дал сходные результаты .
В целом, обнаруженное сходство северных русских с популяциями Северной и
Центральной Европы позволяет выдвинуть гипотезу о сохранении на этих территориях
древнего генофонда Европы, восходящего, возможно, еще к эпохе мезолита. Этот
палеоевропейский генофонд унаследован как северными древнерусскими популяциями, так
и частью финноязычных племен, расселившихся здесь, а также балтоязычными и германоязычными популяциями.
http://www.med-gen.ru/ar/ar_Balanovsky.pdf
Распределение частот гаплогрупп мтДНК у русских в сравнении с другими славянскими популяциями



Russian ethnic history inferred from mitochondrial DNA diversity
Irina Morozova et al.
With the aim of gaining insight into the genetic history of the Russians, we have studied mitochondrial DNA diversity among a number of modern Russian populations. Polymorphisms in mtDNA markers (HVS-I and restriction sites of the coding region) of populations from 14 regions within present-day European Russia were investigated. Based on analysis of the mitochondrial gene pool geographic structure, we have identified three different elements in it and a vast “intermediate” zone between them. The analysis of the genetic distances from these elements to the European ethnic groups revealed the main causes of the Russian mitochondrial gene pool differentiation. The investigation of this pattern in historic perspective showed that the structure of the mitochondrial gene pool of the present-day Russians largely conforms to the tribal structure of the medieval Slavs who laid the foundation of modern Russians.Our results indicate that the formation of the genetic diversity currently observed among Russians can be traced to the second half of the first millennium A.D., the time of the colonization of the East European Plain by the Slavic tribes. Patterns of diversity are explained by both the impact of the native population of the East European Plain and by genetic differences among the early Slavs.
Link
Генетически наиболее близки по мтДНК оказались оказались норвежцы и немцы, в кластер вошли также австрийцы, швейцарцы, поляки, боснийцы, литовцы, ирландцы, шотландцы.  Родственники русских по Аутосомам - не половым хромосомам.
Родственники русских по Аутосомам - не половым хромосомам.



http://www.med-gen.ru/ar/ar_PocheshovaEA.doc
Биологические, археологические и культурологические доказательства палеоазиатского происхождения северных монголоидов, европеоидов и американских индейцев
Ученые России обнаружили европеоидные частоты генов белков и ферментов крови (А.Ф.Назарова, М.Г.Кузнецова. Генетическая структура популяций алтайцев. Доклады РАН, 1993, 333, 3, 405-409.) в семи популяциях алтайцев. Вычисление генетических расстояний 11 человеческих популяций Европы, Азии и Америки, а именно саамов (лопарей), ненцев, нганасан, эвенков, якутов, монголов, алтайцев, русских, финнов, немцев и американских индейцев по частотам 28 аллелей белков, ферментов и групп крови показало определенную близость по наследственным признакам европеоидов и северных монголоидов (Табл.ниже).
 Отечественные генетики вычислили матрицу генетических расстояний 55 человеческих популяций, принадлежащих к четырем большим человеческим расам, и обитающим в Европе, Азии, Америке, Африке и Океании (А.Ф.Назарова, С.М.Алхутов. Генетические расстояния 55 человеческих популяций Европы, Азии, Америки, Африки и Океании по 28 аллелям 12 локусов белков, ферментов и групп крови. В кн.: Эволюция человеческих популяций, М., Полимедиа, 2000, с.14-20.). По данным этой матрицы мы построили эволюционную дендрограмму этих 55 популяций (Рис. 1). Из этой дендрограммы видно, что первыми отделяются, американские индейцы, затем саамы.
Отечественные генетики вычислили матрицу генетических расстояний 55 человеческих популяций, принадлежащих к четырем большим человеческим расам, и обитающим в Европе, Азии, Америке, Африке и Океании (А.Ф.Назарова, С.М.Алхутов. Генетические расстояния 55 человеческих популяций Европы, Азии, Америки, Африки и Океании по 28 аллелям 12 локусов белков, ферментов и групп крови. В кн.: Эволюция человеческих популяций, М., Полимедиа, 2000, с.14-20.). По данным этой матрицы мы построили эволюционную дендрограмму этих 55 популяций (Рис. 1). Из этой дендрограммы видно, что первыми отделяются, американские индейцы, затем саамы.

http://www.trinitas.ru/rus/doc/0211/002a/1087-naz.pdf
Ради интереса: Российский антрополог Т.В Томашевич обнаружила градиент распределения частот надглазничных каналов черепа человека (Табл.ниже)

, (Томашевич Т.В. Закономерности распределение частот надглазничных каналов черепа человека. Вопр. Антропол,1988, вып. 80, с.119-128.).
В надглазничных каналах проходят надглазничные и надблоковые артерий и вен — ветвей глазничной артерии. В исключительных случаях в них проходят надглазничная и надблоковая ветви лобного нерва. Как видно из данных Табл. , у саамов, американских индейцев, а также северных монголоидов - якутов, юкагиров, эвенков, чукчей, эскимосов наиболее высокая частота встречаемости надглазничных каналов, что возможно, вызвано их палеоазиатским происхождением. У кетов, юкагиров, русских, манси, осетинов, армян частота надглазничных каналов очень близка и варьирует в пределах 30-38 %, это может быть подтверждением их общего азиатского происхождения. У монголоидов Восточной Азии – японцев, корейцев, китайцев, а также у монголов частота надглазничных каналов 43-48%. Интересно, что у древних египтян, этрусков, древних славян (полян киевских) и индийцев Уттар Прадеш частота каналов практически одинакова и составляет 25-28%, чуть выше она у словен новгород-ских; это также может быть связано, с происхождением этих народов из древней палеоазиатской популяции.
Итак, родственники русских по всем трём показателям (хромосомы половые и неполовые), - поляки. Русские и Поляки прямые потомки легендарных вандалов, об этом сказано даже в Википедии.
http://ru.wikipedia.org/wiki/%D0%92%D0%B0%D0%BD%D0%B4%D0%B0%D0%BB%D1%8B

Рюрик происходил из севера Германии, из Пруссии, из народов Ругов или Вандалов. На тот момент, 9 век вряд ли вандалы с ругами уже отличались. Т.е он вандал, скорей всего из города Любека с юга Балтики. Вот карта той славянской Германии
Север Германии:

А вот современная карта от Гугла:

Если посмотреть на карту Германии, то в ее северной части можно заметить много названий славянского корня. Городки: Рерик, Руссов, Руггов...С точки зрения генетики, Русские находятся между немцами и поляками, и приходяться как бы на некогда славянские земли Шлезвиг-Гольштейн (земля вандалов силингов) и Верхняя Померания (земля поморов мореплателей).


 Источник:Swedish genetic substructure + something interesting about the Vologda Russians
Источник:Swedish genetic substructure + something interesting about the Vologda Russians
http://eurogenes.blogspot.ru/2011/02/swedish-genetic-substructure-something.html
То есть до сих пор сохраняется генетическое родсто с онемечеными славянами. А теперь о Родстве русских, лужичан, поляков. Русские, Лужицкие сербы и Поляки генетически одна популяция:
http://eurogenes.blogspot.com/2011/05/german-sorbs-genetically-closer-to.html

Популяционная генетика подтверждает полное родство русских с народами южной Балтики, с поляками и лужичанами,- менее выраженное родство с эстонцами, латышами, литовцами. См:
http://svpressa.ru/society/article/60385/
http://www.dzd.ee/449810/analiz-dnk-u-latyshej-litovcev-jestoncev-i-russkih-obwie-geny/

Как уже было продемонстрировано подтверждает только частичное родство с народами северной Балтики: норвежцами по женской линии!.
Тесты:
 Джон Новембер и его коллеги из Калифорнийского университета в Лос-Анджелесе провели исследование, благодаря которому теперь можно с точностью до нескольких сот километров определить место происхождения любого европейца только на основе его ДНК.
Джон Новембер и его коллеги из Калифорнийского университета в Лос-Анджелесе провели исследование, благодаря которому теперь можно с точностью до нескольких сот километров определить место происхождения любого европейца только на основе его ДНК.

Ещё карта Джона Новембера
Novembre's team's map, showing predicted locations for each of 1,387 individuals based on their DNA.

http://www.newscientist.com/article/dn14

http://dienekes.blogspot.com/2009/02/auton-et-al-on-global-distribution-of.html
Русские, Белорусские Славяне, Прибалты и скандинавы наиболее однообразны, после британцев. Генетик и антрополог Dienekes подвёл итоги проекта по обработке SNP-материала и опубликовал дополнение, которого не хватало в недавнем кластером анализе, — наглядные диаграммы главных компонент: от первой до пятой.


 Как видно, первая главная компонента отделяет Западную Евразию от Индии, вторая задаёт клинальный градиент между северными европейцами и средиземноморцами, третья выделяет из массива Западной Евразии североафриканское влияние, четвёртая — ашкеназов, пятая — финнов.Обращает на себя внимание плотная группировка русских со скандинавами, скандинавов — с немцами и ирландцами, половины немцев — с жителями Балкан, и относительная близость испанцев к северным европейцам. Показательно также, что по второму компоненту индусы находятся строго на уровне северных европейцев, не смещаясь по средиземноморской оси в сторону турок, армян и прочих греков.
Как видно, первая главная компонента отделяет Западную Евразию от Индии, вторая задаёт клинальный градиент между северными европейцами и средиземноморцами, третья выделяет из массива Западной Евразии североафриканское влияние, четвёртая — ашкеназов, пятая — финнов.Обращает на себя внимание плотная группировка русских со скандинавами, скандинавов — с немцами и ирландцами, половины немцев — с жителями Балкан, и относительная близость испанцев к северным европейцам. Показательно также, что по второму компоненту индусы находятся строго на уровне северных европейцев, не смещаясь по средиземноморской оси в сторону турок, армян и прочих греков.
Для создания генетического портрета населения Архангельской области использована вся доступная нам информация о полиморфизме четырех систем признаков - Y хромосомы, митохондриальной ДНК (мтДНК), аутосомных ДНК маркеров и квазигенетических маркеров - фамилий (табл. 1, рис. 1). Использовались как архивные и опубликованные данные лаборатории популяционной генетики человека МГНЦ РАМН, так и опубликованные данные других авторов. В контексте данной работы термины «генофонд населения Архангельской области» и «генофонд Русского Севера» использованы как синонимы. ДНК коллекции лаборатории популяционной генетики человека Медико-генетического научного центра РАМН (далее сокращенно обозначается ЛПГЧ), представляющие основной исторический ареал русского народа, генотипировались О.П. Балановским по мтДНК и частично по Y хромосоме в Эстонском биоцентре РАМН (рук. Р. Виллемс), остальная часть данных по Y хромосоме и данные по аутосомным ДНК маркерам получены в ЛПГЧ (табл. 1).



http://eurogenes.blogspot.ru/2012/04/so-whos-most-european-of-us-all.html
У вологодцев насчитали самый большой % северных генов, у русских из центральных областей он меньше
 http://eurogenes.blogspot.com.au/2012/09/they-had-blond-hair-and-light-eyes-and.html
http://eurogenes.blogspot.com.au/2012/09/they-had-blond-hair-and-light-eyes-and.html
Ещё два типа европеоидов - средние и южные - представлены в разных переходных вариантах носителями гаплогруппы R1a1 (типичные представители - большинство русских, поляков, украинцев, белорусов, словаков, чехов, венгров) и R1b (типичные представители - ирландцы, испанцы, баски, французы, датчане, половина немцев, две трети итальянцев). Очевидно, что во втором типе перемешаны средние и южные европеоиды.
Список родственников русских:
Лужичане, Поляки, Украинцы, Белорусы, Венгры, Чехи, Словаки, Латыши, Литовцы, Киргизы, Алтайцы, Таджики, Ишкашим, Белуджи, Пенджабцы, Пуштуны, Талыши, Норвежцы,Немцы, Австрийцы, Швейцарцы, Поляки, Боснийцы, Литовцы, Ирландцы, Шотландцы, Иранцы, Коми, Чуваши, Удмурты, Ненцы,Осетины, Азербайджанцы.То что русские родственники киргизов, таджиков, можно не сомневаться - вот например Дерево родства 61 евразийской популяции основанное Y-хромосоме. :))

Смотри
http://www.pnas.org/content/98/18/10244.full
http://ru.wikipedia.org/wiki/%C0%ED%F2%F0%EE%EF%EE%EB%EE%E3%E8%FF_%F0%F3%F1%F1%EA%E8%F5

Различные фенотипы цвета глаз (a-d). Людей с голубыми глазами без единого пятнышка (a) относят к генотипу rs12913832 G/G, в то время как всех остальных (b-d) к генотипу rs12913832 A/G (фото Københavns Universitet).
Команда учёных под руководством доцента Ханса Эиберга (Hans Eiberg) из университета Копенгагена (Københavns Universitet) изучила мутацию, которая произошла около 6-10 тысяч лет назад, и обнаружила, что у ныне живущих людей с голубыми глазами, вероятно, был общий предок.
Произошедшая около 10 тысяч лет назад мутация, которая затронула ген OCA2, привела к тому, что производство меланина в радужной оболочке глаза было ограничено (и коричневая окраска была «разбавлена» до голубой).
Если бы OCA2 был бы полностью выключен, человек стал бы альбиносом, так как прекратилась бы выработка меланина в волосах, глазах и коже. В данном случае изменения привели лишь к частичному ограничению его производства организмом.
И хотя цвет глаз определяется различными параметрами, а не только количеством меланина, у голубоглазых его содержание в радужке значительно меньше, чем у всех остальных людей.
Изучая ДНК митохондрий, учёные установили, что у всех людей с голубым цветом глаз присутствует незначительное изменение в определённом месте двойной спирали. В то время как у кареглазых нет какой-то одной характерной «точки», их вариации в строении ДНК (в той её области, что отвечает за процессы, происходящие с меланином в организме) более обширны.
Из этого учёные и сделали вывод о том, что у всех голубоглазых был один общий предок, у которого и произошла данная мутация.
http://www.membrana.ru/particle/12269

Региональное расположение кластеров:
— жёлтый — Западная Азия;
— розовый — Средиземноморье;
— зелёный — Северная Европа;
— синий — Южная Балтика;
— голубой — Волга-Урал;
— красный — Сибирь.
Таблица расстояний между кластерами:

Доля зелёного «северо-европейского» компонента достигает максимума в Скандинавии и Финляндии, в то время как среди остальных популяций он ниже в полтора раза, а у чувашей отсутствует полностью. Синий «южно-балтийский» компонент распределён схожим образом, но менее широко: он встречается у славян, балтов и венгров, но практически отсутствует в Скандинавии, среди финнов и чувашей. Голубой компонент «Волга-Урал» доминирует у чувашей, высок у русских из Вологды, прибалтийских и поволжских финнов, ниже у остальных русских и тонко распределён по другим северо-восточным европейцам. Расстояния от волго-уральского кластера до двух других меньше (0,0017 и 0,0020), чем между южно-балтийским и северо-европейским кластерами (0,0025).
Все три кластера ассоциируются с Северо-Восточной Европой, явно имеют общее происхождение и отражают процесс формирования облика современного северо-европейского населения. Тёмно синий «южно-балтийский» компонент здесь отчётливо связан с расселением славян и балтов по северо-востоку континентальной Европы (культуры шнуровой керамики?), нёсших поровну зелёного и характерного синего. Голубой «волго-уральский» компонент — это, вероятно, архаичный европеоидный субстрат, сохранивший свои позиции в северо-восточных областях континента как в качестве предков как не связанного с финнами населения (поздние восточные потомкисвидерской культуры: нарвская, верхневолжская, неманская?), так и предков прибалтийских финнов (финноязычные народы формировались на европеоидной основе). В континентальной части это население испытало влияние балтов/славян, местами с последующим возвратом к финской культуре: структура поволжских финнов наполовину южно-балтийская, наполовину — финская.
Что касается зелёного «северо-европейского» кластера, можно согласиться с Дэвидом, что в него вошли все те автохтонные в Северной Европе аллели, которые не попали в «южно-балтийский» и «волго-уральский» кластеры. Он не демонстрирует корреляции с германскими языками и не выделяется как изолированный от двух своих северных братьев компонент, но в целом показывает бóльшую близость к более архаичному «волго-уральскому» кластеру. Вероятно, он охватывает всё то северо-европеоидное население, которое широко расселилось по северу Европы к моменту начала экспансии предков балтов и славян, но не было затронуто влиянием последних (в Скандинавии — потомки аренсбургской культуры и последующих?).
Если рассматривать эти данные генетики с точки зрения антропологии и археологии, то они не открывают ничего нового, укладываются в непротиворечивую, давно известную картину. В действительности, они рассказывают историю Европы с собственной точки зрения и акцентируют внимание на том, что важно для популяционной генетики как таковой. В частности, мы видим отчётливое указание, что к началу формирования на континенте балтов и славян в Скандинавии и Финляндии уже жило родственное им северо-европеоидное население, а у них обоих были ещё одни, более древние предки.
http://drfaust-spb.livejournal.com/261396.html
Аутосомные ДНК о происхождении славян и балтов
Вернёмся к последней диаграмме структурного анализа ДНК северо-востока Европы, чтобы извлечь из неё ещё не извлечённые следствия.
Немного общих мест. Популяционная генетика примечательна тем, что рассказывает историю народов на языке непосредственно данных о населении, без обращения к производным от населения: археологии и лингвистике. Особенность аутосомных ДНК в том, что они показывают, чьим именно потомком является тот или иной современный человек, — за счёт равного наследования структуры ДНК от обоих родителей из поколения в поколение (в отличие от данных однородительских маркёров, которые позволяют выделить дальних предков человека по материнской и отцовской линиям). Другими словами, результат структурного анализа аутосомных ДНК указывает, откуда пришли предки человека, пропорционально их удельному весу.

В проекте Dodecad Ancestry Project был проведен fastIBD анализ по популяциям Центральной и Восточной Европы. Синими линиями я выделил Восточно-Европейский кластер, куда попали восточные славяне: русские, белорусы, украинцы. Зеленый квадрат выделенный красными линиями это и есть русские.

Структура русского генофонда

Как показывают результаты анализа ДНК, генетическая преемственность между древними и современными народами соблюдается отнюдь не всегда, ведь одни народы могли сменяться другими. Например, генофонды неолитического и нынешнего населения Прибайкалья имеют мало общего . На возможную смену этносов в некоторых европейских регионах - Тоскании (Италия) и Стране басков (Испания) - указывают результаты генетических исследований древнего и современного населения . Более того, уже имеются примеры существенных противоречий между наблюдаемыми и ожидаемыми распределениями маркеров мтДНК в палеопопуляциях. Так, у неолитического населения Страны басков не обнаружена митохондриальная группа V. А ведь именно ей придается большое значение, когда, основываясь на распределении групп мтДНК у современных европейцев, моделируют процесс послеледникового заселения Европы выходцами из пиренейского "убежища" .
Выявлено и еще одно серьезное несоответствие. У неолитического населения Центральной Европы, ведущего происхождение от "первых европейских фермеров" (носителей переднеазиатской технологии агрокультуры), митохондриальная группа N1a встречается с неожиданно высокой частотой. Современным же европейцам эта группа практически не свойственна и потому никак нельзя было предсказать ее высокую распространенность среди неолитического населения .
Приведенные примеры показывают, насколько важна проверка этноисторических моделей, если выявленное генетическое разнообразие современных народов переносится в их прошлое. Выходит, экстраполяции не всегда надежны. Каким был генофонд далеких предков, безошибочно можно судить по результатам исследований ДНК непосредственно древнего населения. И такой анализ вполне возможен, ведь археологами найдено множество останков неолитического времени, пригодных для выделения ДНК.
Из работы ХРОМОВОЙ НАТАЛЬИ АНАТОЛЬЕВНЫ;
2. Проанализировать генетическое родство исследованных популяций, как между собой, так и с некоторыми народами Европы.
3. На примере восточных славян изучить способность системы генов HLA отражать геногеографию этнических групп и возможность использования этой системы в популяционных исследованиях.


1. Самыми частыми аллельными вариантами DRB1 гена для всех изученных популяций являются: DRB1*07, *11, *13, *15, что соответствует западно-евразийскому типу распределения.
2. При попарном сравнении распределения специфичностей гена DRB1 между восточными украинцами и русскими выявлены достоверные различия в частотном содержании DRB1*11 и DRB1*16.
3. При попарном сравнении распределения аллелей гена DRB1 выявлено, что западные украинцы достоверно отличаются от русских частотным содержанием DRB1*11.
4. Сравнительный анализ представленных данных отмечает, что у всех популяций часто встречаемыми являются следующие трехлокусные гаплотипы: DRB1*11-DQA1*0501-DQB1*0301, DRB1*15-DQA1*0102-DQB1*0602-8, DRB1*01-DQA1*0101-DQB1*0501, DRB1*04-DQA1*0301-DQB1*0302 и DRB1*07-DQA1*0201-DQB1*0201
5. Отличительной особенностью является отсутствие или низкие частоты гаплотипов: DRB1*09-DQA1*0301-DQB1*0303 и DRB1*12-DQA1*0501-DQB1*0301
6. При попарном сравнении частот распределения трехлокусных гаплотипов (DRB1-DQA1-DQB1) восточных украинцев с русскими было установлено, что достоверными являются различия в частотном содержании DRB1*11-DQA1*0501-DQB1*0301 и DRB1*16-DQA1*0102-DQB1*0502/4.
7. Результаты многомерного статистического анализа выявили, что изученные популяции образуют два кластера»: «южный», включающий украинцев (Хмельницкая и Львовская области), и «северный», включающий белорусов (Витебская и Брестская области) и русских из Вологодской области.
8. При проведении многомерного кластерного анализа изученных популяций и некоторых народов Европы выявлено, что белорусы из Витебской и Брестской областей и украинцы из Хмельницкой и Львовской областей генетически близки с «центрально-европейскими» популяциями. Русские из Вологодской области сходны по HLA–профилю с «североевропейскими» и «центрально-европейскими» популяциями.
9. На примере восточных славян показано, что, несмотря на влияние естественного направленного отбора, система генов HLA адекватно отражает геногеографию этих этнических групп, и может быть использована в популяционных исследованиях.
Русский генофонд - Новое генетическое исследование русских
Analysis of Y chromosome STR haplotypes in the European part of Russia reveals high diversities but non-significant genetic distances between populations Были исследованы Y-хромосомы 545 русских мужчин из Смоленской, Брянской, Ивановской, Липецкой, Пензенской, Рязанской, Орловской, Тверской, Вологодской, Тамбовской, Архангельской и Новгородской областей, т.е. по сути дела всего ареала образования великороссов.
АЛЛЕЛЬНЫЙ ПОЛИМОРФИЗМ ШЕСТИ МИКРОСАТЕЛЛИТНЫХ ЛОКУСОВ ДНК В ПОПУЛЯЦИЯХ РЕСПУБЛИКИ САХА (ЯКУТИЯ)
Ключевые слова: аутосомные микросателлитные локусы, популяции Якутии, генетическое разнообразие.
ALLELIC POLYMORPHISM OF SIX MICROSATELLITE DNA LOCI IN POPULATIONS OF SAKHA (YAKUTIA), by V. V. Zhirkova, S. A. Fedorova, V. L. Akhmetova, L. A. Zhivotovsky, E. K. Khusnutdinova, Yakut Research Center of Complex Medical Problems, Russian Academy of Medical Sciences, Yakutsk, 677010 Russia
Якутия расположена в северо-восточной части Евразийского континента. На обширной территории, занимающей по площади 1/5 часть территории России, проживает менее 1 млн. человек, представляющих более сотни разных этносов. Среди них большинство составляют русские, якуты, эвенки, эвены, юкагиры и долганы. В настоящее время из представителей коренного населения Якутии лишь у якутов определены частоты 10 аутосомны STR локусов . Более интенсивно полиморфизм ауто сомных диаллельных и минисателлитных маркеров , мтДНК Y-хромосомы также изучали ранее лишь у якутов как наиболее многочисленного этноса Сибири. Оказалось, что популяция якутов имеет более низкие показатели генетического разнообразия, чем другие тюркоязычные этносы и резко отличается от других этносов по своим генетическим характеристикам. Учитывая это, изучение полиморфизма микросателлитны STR-локусов в популяциях коренного населения Якутии — русских, якутов, юкагиров, долган и эвенков, представляет большой интерес как прикладной — для развития популяционных баз данных, так и фундаментальный — для выявления особенностей структуры генофонда отдельных этносов
Цель настоящей работы — изучение полиморфизма шести аутосомных микросателлитных локусо (D3S1358, D16S539, THOI, D8S1179, LPL, HUM-vWFII)для создания референсной базы популяционно-генетических данных в семи популяциях коренного населения Якутии, необходимой при вычислении вероятностной статистики.
ЭКСПЕРИМЕНТАЛЬНАЯ ЧАСТЬ
В работе использованы 317 образцов ДНК коренных жителей Республики Саха (Якутия). Коллекция составлена из материала Банка ДНК Отдела молекулярной генетики ЯНЦ КМП РАМН, собранного в экспедиционных выездах 2001—2006 гг. В выборку вошли неродственные индивиды, этническую принадлежность которых учитывали до третьего поколения. Всего проанализированы семь популяций: центральные якуты (n = 53), населяющие Лено-Амгинское междуречье (Чурапчинский, Усть-Алданский, Хангаласский, Амгинский, Мегино- Кангаласский, Намский улусы), вилюйские якуты (n = 49), проживающие в бассейне реки Вилюй (Сунтарский, Нюрбинский, Вилюйский, Верхне - вилюйский улусы), северные якуты (n = 53) (Верхоянский, Момский, Абыйский, Среднеколымский улусы), эвенки Жиганского, Усть-Майского, Оленекского улусов (n= 49), юкагиры Верхнеколымского и Нижнеколымского улусов ( n= 53), долганы Ана- барского улуса (n = 13), русскиe (n = 47) — жители населенных пунктов Мирнинского, Ленского, Алданского, Нерюнгринского улусов.
Суммарную ДНК выделяли из лимфоцитов периферической крови методом фенол-хлороформной экстракции с использованием протеиназы К. Матричные препараты ДНК амплифицировали методом ПЦР с помощью реагентов ТАПОТИЛИ (Государственный научный центр Российской Федерации “ГосНИИгенетика”, Москва). Амплификацию проводили на приборе Терцик (“ДНК-технология”, Россия). Продукты амплификации разделяли с помощью электрофореза в 10%-ном полиакриламидном геле с последующим окрашиванием бромистым этидием и серебром и визуализировали в УФ- свете системы гель-видеодокументации (“Vilber Lourmat”, Франция).
Статистический анализ данных (частоты аллелей и их ошибки, фактическая и теоретическая гетерозиготность, соответствие распределения генотипов равновесию Харди—Вайнберга) проводили с помощью пакета компьютерных програм Genepop (versio 3.3, 2001). Ожидаемую гетерозиготность и параметры информативности локусов: вероятность случайного совпадения генотипов двух неродственных индивидо (MF),вероятность дискриминации генотипов двух неродственных индивидо (PD) информативность полиморфизма данного локус (PIC) рассчитывали с помощью компьютерной программ PowerTyperExcel spreadsheet (“Promega” . Достоверность различий в распределении частот аллельных вариантов между популяциями оценивали с помощью точного теста программ RxC Факторный анализ с использованием метода главных компонент проводили с помощью программ POPSTR любезно предоставленно H. Harpending.


Микросателлит THOI расположен в первом интроне гена тирозингидроксилазы, локализованного на коротком плече хромосомы 11. Описано восемь аллелей этого локуса с числом повторяющихся единиц от пяти до 11. В выборках юкагиров выявлено семь аллелей маркер THOI, у русских — шесть аллелей, у долган только четыре, а во всех остальных популяциях — пять аллелей. Из них в популяциях центральных, вилюйских якутов, эвенков и юкагиров наиболее распространен аллель с семью повторами. Согласно, аналогичное преобладание аллеля 7 характерно для популяций бурятов, сойотов, хакасов, чукчей, коряков и хамниганов. В популяциях русских, долган и северных якутов с наибольшей частотой встречается аллель 9.3, характерный также для народов Волго-Уральского региона, Центральной России, Северной Кореи . Индексы наблюдаемой гетерозиготности (Hobs варьировали от 0.740 в популяции северных якутов до 0.880 у эвенков. Среднее значение фактической гетерозиготности в изученных группа (Hobs=0.810) оказалось выше средней теоретической гетерозиготности (Hexp= 0.766).
Микросателлит D8S179 расположен на хромосоме 8q24.1—q24.2. В пяти популяциях обнаружили восемь различных аллелей этого маркера. В выборках юкагиров и долган найдено шесть и пять аллелей соответственно. Во всех популяциях отмечен унимодальное распределение аллелей с максимальной частотой аллеля 13. В целом, наши данные согласуются с результатами, полученными для народов Южной Сибири, Волго-Уральского региона, Центральной России, Северной Кореи, Чукотки и Камчатки, где также наблюдается одномодальное распределение частот аллелей с преобладанием аллеля 13 . Показатель теоретической гетерозиготности в среднем для семи популяций коренного населения Якутии составил Hexp= 0.650.
Всего в изученных нами популяциях выявлено 45 аллелей. В отдельных выборках число аллелей варьирует от 26 (у долган) до 40 (у русских). Анализ шести судебных локусов показал высокий уровень разнообразия среди популяций Якутии. В изученных выборках обнаружены по семь—восемь аллелей четырех из шести маркеров (восемь D16S539, D8S1179 и семь HUMvWFII и THOI).
Во всех популяциях не найдено существенных отклонений от равновесия Харди—Вайнберга.При попарном сравнении частот аллелей маркеров D3S1358, LPL, THOI, D8S1179 HUMvWFI становлены достоверные различия между русскими и остальными популяциями. Анализ гетерогенности микросателлитных маркеров D16S539, HUMvWFI выявил достоверные отличия долган от всех остальных популяций. Показано, что по распределению частот аллелей микросателлит D8S1179 популяция юкагиров достоверно отличается от всех остальных популяций.
По уровню молекулярного разнообразия все локусы относятся к средневариабельным (число наблюдаемых аллелей 5 ≤ V ≤ 8). В зависимости от значения PIC высокоинформативными оказались маркеры D16S539, D3S1358, THOI, D8S1179, HUMvWFII (PIC> 0.5). Маркер LPL оценен как умеренно информативный (0.5 > PIC> 0.25) (табл. 3)
 Описанное выше высокое внутрипопуляционное разнообразие отражается в значительном дискриминирующем потенциале этой полиморфной системы. Высокие значения индекса дискриминации (PD), изменяющиеся в диапазоне от 0.90 до 0.91, определяются в трех из шести локусов — D3S1358, THOI, D16S539. В суммарной выборке мы не встретили двух индивидов с одинаковым мультилокусным генотипом по шести маркерам. Дискриминирующий потенциал по совокупности шести локусов изменяется от 2.89 х 10-4 долганов до 1.32 х 10-6 русских. Другими словами, случайно выбрать из популяции два одинаковых мультилокусных генотипа у неродственных индивидов можно лишь в одном случае из шести миллиардов. Это означает, что фактически анализируемые маркеры подходят по всемсуществующим критериям для решения идентификационных задач.
Описанное выше высокое внутрипопуляционное разнообразие отражается в значительном дискриминирующем потенциале этой полиморфной системы. Высокие значения индекса дискриминации (PD), изменяющиеся в диапазоне от 0.90 до 0.91, определяются в трех из шести локусов — D3S1358, THOI, D16S539. В суммарной выборке мы не встретили двух индивидов с одинаковым мультилокусным генотипом по шести маркерам. Дискриминирующий потенциал по совокупности шести локусов изменяется от 2.89 х 10-4 долганов до 1.32 х 10-6 русских. Другими словами, случайно выбрать из популяции два одинаковых мультилокусных генотипа у неродственных индивидов можно лишь в одном случае из шести миллиардов. Это означает, что фактически анализируемые маркеры подходят по всемсуществующим критериям для решения идентификационных задач.
Далее с использованием частот аллелей микро- сателлитных локусов сравнивали популяции Якутии и других регионов (Южная Сибирь, ВолгоУральский регион, Центральная Россия, Северная Корея, Чукотка и Камчатка) . В пространстве двух главных компонент, описывающих 40% вариабельности аллельных частот, рассматриваемые популяции (рис. 2)

четко разделяются по первой компоненте на два кластера — первый образован европеоидными популяциями и тофаларами, второй — монголоидными популяциями Азии. В этом случае аллели, имеющие более высокую частоту в европеоидных популяциях, — это аллель 6 локуса THOI, аллель 18 локуса D3S1358, аллель 14 локуса D8S1179. Монголоидные популяции характеризуются высокими частотами аллеля 9 локуса D16S539, аллеля 15 локуса D3S1358, аллеля 7 локуса THOI, аллеля 13 локуса D8S1179. По вариабельности частот микросателлитных локусов популяции Якутии группируются вместе с популяциями Южной Сибири, отдалены от популяций Чукотки и Камчатки, что соответствует данным анализа мтДНК.
К ПРОБЛЕМЕ ПРОИСХОЖДЕНИЯ МОНГОЛОИДНОГО КОМПОНЕНТА МИТОХОНДРИАЛЬНОГО ГЕНОФОНДА СЛАВЯН





В табл. 2 показаны результаты иерархического анализа изменчивости мтДНК в группах популяций. Видно, что при группировании популяций в соответствии с их этнолингвистической характеристикой (группа русских, группа поляков и группа балтского и угро-финского происхождения) межгрупповые различия очень низки и недостоверны. Однако выделение отдельной группы популяций, включающей северо-западных русских (Великий Новгород и Псков), северо-восточных поляков (Сувал-ки) и популяции балтского и угро-финского происхождения, приводит к достоверному увеличению степени межгрупповой дифференциации (0.15%). Еще большие различия наблюдаются, если отдельная группа представлена только северо-западными русскими и северо-восточными поляками. Это свидетельствует о своеобразии генетической структуры этих популяций по отношению как к остальным славянским популяциям (русским и полякам; 0.52%), так и к популяциям балтско-го и угро-финского происхождения (0.76%). Анализ генофондов северо-западных русских и северо-восточных поляков в сравнении с их непосредственными соседями - литовцами и эстонцами - также показывает достоверные различия между ними (0.73%). Таким образом, генетическое своеобразие русского населения Северо-Западного региона России и Северо-Восточной Польши не может быть объяснено исключительно лишь их сходством с балтским и угро-финским населением, а является следствием появления уникальной генетической композиции, сложившейся, по-всей видимости, в результате взаимодействия трех генофондов - славянского, балтского и угро-финского. Анализ структуры митохон-дриального генофонда популяций Великого Новгорода, Пскова и Сувалок показал наличие лишь одного генетического компонента -гаплогруппы U5a, которая распространена в этих популяциях с более высокой частотой (в среднем, 16%), чем в соседних славянских, балтских и угро-финских популяциях, где ее частота, в среднем, составляет 7%. Интересной особенностью генофондов популяций Великого Новгорода, Пскова и Сувалок является также присутствие (в среднем, с частотой 2.2%) гаплотипов митохондриальных групп R1 и R2, которые крайне редки в соседних популяциях.
Генетические параллели между населением Северо-Западной России и Северо-Восточной Польши











 На самом деле у генетиков получается прямо противоположное
На самом деле у генетиков получается прямо противоположное
Какой бы признак мы ни взяли, видим: русские – типичные европейцы, а азиатское завоевание оставило след в их истории, но не в генофонде. Не беремся говорить о России и русских в культурном, историческом, гуманитарном смысле. Однако биологически их генофонд не является промежуточным между европейским и азиатским, он – типичный европейский. Уточним: самый восточный из этого ряда, стоящий «на передовой». Отдельные азиатские влияния прослеживаются в нём больше, чем в западных соседях. Но базовый, главный вывод, который следует из проведенного исследования – практически полное отсутствие в русском генофонде монголоидного вклада
Елена БАЛАНОВСКАЯ, Олег БАЛАНОВСКИЙ
Русский генофонд: свидетельства «очевидцев»
http://www.den-za-dnem.ru/page.php?article=322
Отличия русского генофонда (почти полностью европейского) от монгольского (почти полностью центрально-азиатского) действительно велики – этo как бы два разных мира. Но если говорить не о монголах, а о татарах, с которыми чаще всего и имели дело русские княжества, то отличия их генофонда от русского не такие уж и большие. Различия между русским и татарским генофондом хотя и есть, они вовсе не колоссальные – русский полностью европейский, а татарский – по большей части европейский. Мы сделали вывод, и он был многократно подтвержден, что в русском генофонде практически нет следов из Азии, из-за Урала.Олег БАЛАНОВСКИЙ
Сенсационное открытие ученых: Раскрыта тайна русского генофонд
http://www.kp.ru/daily/24065/305831/
Доля восточно-евразийских гаплогрупп в русском населении составила лишь 2%. То есть очень малую величину. Почти столь же малую, что и в генофонде поляков (1,5%) или севера Европы, где уже точно никаких «татаро-монгольских вкладов» не было (у норвежцев 0,6%, у исландцев 0,7%, у карел 4,8% и т.д.). Средняя «фоновая» частота восточно-евразийских гаплогрупп в Европе равна 3,6%. То есть в русском генофонде (2,0%) она даже меньше, чем «средняя по Европе», поэтому монголоидный компонент у русских оказывается не просто нулевым, но даже с отрицательным знаком. Итак, мы не видим последствий монгольского нашествия в русском генофонде – или тогда должны видеть эти последствия и в польском, и в норвежском, и в карельском, и в других генофондах Европы
Е.В. Балановская, О.П. Балановский. Русский генофонд на Русской равнине. М., 2007. С. 296 Что же касается второй из крупных миграций, связанных с монгольским завоеванием средневековых русских княжеств, то ее генетические следы обнаружить не удается. И вновь этот вывод взаимно подтверждается анализом и мтДНК, и Y-хромосомы, и данными антропологии. Например, суммарная частота восточно-евразийских гаплогрупп мтДНК в русских популяциях не достигает и 2%: эта же частота характерна и для западноевропейских народов. Для Y-хромосомы типичным «монгольским» маркером является гаплогруппа С (ее носителем был, как считается, Чингисхан, – эта гаплогруппа является самой частой у монголов и родственных им народов). Однако в русских популяциях гаплогруппа С практически не встречена (частота не достигает 1%, т. е. с формально-генетических позиций полиморфизм по этому признаку в русских популяциях отсутствует, и русское население может считаться полностью «генетически европейским»).Е.В. Балановская, О.П. Балановский
ГЕНЕТИЧЕСКИЕ СЛЕДЫ ИСТОРИЧЕСКИХ И ДОИСТОРИЧЕСКИХ МИГРАЦИЙ: КОНТИНЕНТЫ, РЕГИОНЫ, НАРОДЫ
http://www.bionet.nsc.ru/vogis/pict_pdf/2009/2009_2/20.pdf
Иностранный источник по данной тематике...
Данные генетики об азиатском влиянии на русских

Данные генетики об азиатском влиянии на русских
Исследование взаимовлияний европейских и азиатских популяций по Х-хромосоме вновь подтверждает факт, установленный более ранними исследованиями: азиатское влияние на русский генофонд крайне незначительно. На приведенной ниже таблице представлено соотношение европейских (слева) и азиатских (справа) компонентов в ряде европейских и азиатских популяций. Из нее следует, что незначительная азиатская примесь имеется у всех европейских популяций, причем у русских она примерно на том же уровне, что и у германских народов - голландцев, немцев и шведов
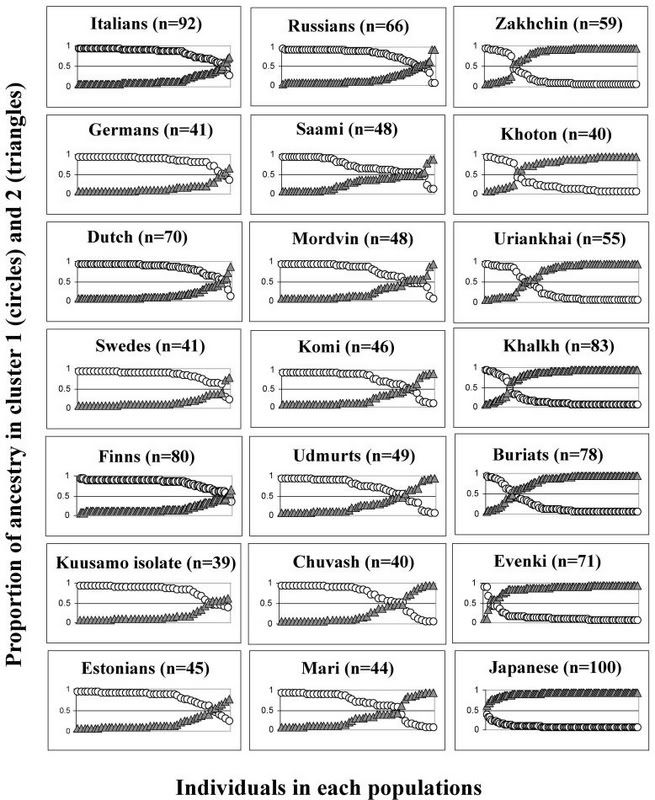
С исследованием можно ознакомиться здесь:
С исследованием можно ознакомиться здесь:
Карта генетических расстояний между народами мира.
В мировом контексте европейцы генетически – одна семья.
Origins of Mongoloid mtDNA in Slavs
A very interesting article which addresses the issue of the Mongoloid mitochondrial component in Slavs. I like this kind of paper that looks at the minor features of populations, rather than their broad general characteristics that are well known by now, and indeed were known even before genetics came into the picture.
From the paper:
Russian Journal of Genetics, 10.1007/s11177-008-3016-9
On the origin of Mongoloid component in the mitochondrial gene pool of Slavs
B. A. Malyarchuk, M. A. Perkova and M. V. Derenko
Abstract The data on mitochondrial DNA (mtDNA) restriction polymorphism in Czech population (n = 279) are presented. It was demonstrated that in terms of their structure, mitochondrial gene pools of Czechs and other Slavic populations (Russians, Poles, Slovenians, and Bosnians) were practically indistinguishable. In Czechs, the frequency of eastern-Eurasian (Mongoloid) mtDNA lineages constituted 1.8%. The spread of eastern-Eurasian mtDNA lineages belonging to different ethnolinguistic groups in the populations of Europe was examined.Frequency variations of these DNA lineages in different Slavic groups was observed, with the range from 1.2 and 1.6% in Southern and Western Slavs, respectively, to 1.3 to 5.2% in Eastern Slavs, the Russian population of Eastern Europe. The highest frequency of Mongoloid component was detected in the mitochondrial gene pools of Russian populations from the Russian North and the Northwestern region of Russia.This finding can be explained in terms of assimilation of northern-European Finno-Ugric populations during the formation of the Russian population of these regions. The origin of Mongoloid component in the gene pools of different groups of Slavs is discussed.
From the paper:
The overwhelming majority of mtDNA clusters, identified in Czechs, were of the western-Eurasian origin.The frequency of eastern-Eurasian (Mongoloid) mtDNA lineages in this population constituted 1.8% (haplogroups A, N9a, and M). African lineage (with the frequency of 0.4%) belonging to haplogroup L2a and marked by the +13803HaeIII variant was also detected.
...
Low frequency of Mongoloid mtDNA variant in Letts and Lithuanians suggests that Mongoloid component was probably not typical of Balto-Slavic protogene pool. Thus, it seems reasonable that accumulation of Mongoloid mtDNA lineages in Slavs and their ancestors was intensified only in the last 4000 years....
The highest frequencies of the Mongoloid component are typical of the Russian populations from Russian Pomor’e and Northwestern region. These populations, however, differ in the mtDNA haplogroup composition. It was established that assimilation of the indigenous pre-Slavic population of Eastern Europe by true Slavs was of great importance to the process of the development of Russian population....
It is suggested that after the decay of Avar Khaganate the populations included into it were assimilated by Slavic tribes [34]. It is thereby suggested that rather high frequencies of eastern-Eurasian mtDNA lineages observed in the gene pools of some populations of Western and Southern Slavs (especially on the territoryof former Avar Khaganate) can be considered as a consequence of the process described.
Concerning the population of Eastern Europe, it should be noted that the forest zone of Eastern Europe was the area of intense population admixture [35]. It seems likely, that formation of the complex of Mongoloid traits happened not later than in Upper Paleolithic. For this reason, it is suggested that East Siberian populations could have much time for migration to Eastern Europe [35]. The number of such migrations still remains unclear, since in the northwest of Eastern Europe Mongoloid component is detected 10 000–8000 years ago; in Dnepr–Donetsk tribes, 7000–6000 years ago, and on the territory of Ivanovo oblast (Sakhtysh), 6000–5000 years ago [35, 36]. The data on mtDNA variation in Russian populations are consistent with anthropological data, since they point to the substantial differences in the frequencies of Mongoloid mtDNA lineages between the Russian populations of the Russian North, Northwest, and the central/southern regions of the European part of Russia (Table 3). We hope that future investigations would provide
the possibility of analyzing the chronology of the development of the Mongoloid component diversity in the gene pools of Russians and other Slavic populations.
Russian Journal of Genetics, 10.1007/s11177-008-3016-9
On the origin of Mongoloid component in the mitochondrial gene pool of Slavs
B. A. Malyarchuk, M. A. Perkova and M. V. Derenko
Abstract The data on mitochondrial DNA (mtDNA) restriction polymorphism in Czech population (n = 279) are presented. It was demonstrated that in terms of their structure, mitochondrial gene pools of Czechs and other Slavic populations (Russians, Poles, Slovenians, and Bosnians) were practically indistinguishable. In Czechs, the frequency of eastern-Eurasian (Mongoloid) mtDNA lineages constituted 1.8%. The spread of eastern-Eurasian mtDNA lineages belonging to different ethnolinguistic groups in the populations of Europe was examined.Frequency variations of these DNA lineages in different Slavic groups was observed, with the range from 1.2 and 1.6% in Southern and Western Slavs, respectively, to 1.3 to 5.2% in Eastern Slavs, the Russian population of Eastern Europe. The highest frequency of Mongoloid component was detected in the mitochondrial gene pools of Russian populations from the Russian North and the Northwestern region of Russia.This finding can be explained in terms of assimilation of northern-European Finno-Ugric populations during the formation of the Russian population of these regions. The origin of Mongoloid component in the gene pools of different groups of Slavs is discussed.
Кое что о русском дворянстве....Которое вроде было сплошь немцы и татары, а каким оно было на самом деле?
Так по данным приводимым историком Загоскиным Н.Р. (1851 - 1912) в среде русского дворянства было происходивших от Рюрика 168 родов и 43 рода другого русского происхождения. Но одновременно от татар происходили 120 родов (напр. Аксаковы, Бунины, Корсаковы, Салтыковы, Юсуповы и др.) и 16 родов другого восточного происхождения, 223 рода польско-литовского происхождения и более 120 другого западно -европейского происхождения.
Источник здесь.
http://his95.narod.ru/zam6_3.htm
И что же мы имеем фактически? Больше всего было не татарских и немецких, а польско-литовских родов.
Так по данным приводимым историком Загоскиным Н.Р. (1851 - 1912) в среде русского дворянства было происходивших от Рюрика 168 родов и 43 рода другого русского происхождения. Но одновременно от татар происходили 120 родов (напр. Аксаковы, Бунины, Корсаковы, Салтыковы, Юсуповы и др.) и 16 родов другого восточного происхождения, 223 рода польско-литовского происхождения и более 120 другого западно -европейского происхождения.
Источник здесь.
http://his95.narod.ru/zam6_3.htm
И что же мы имеем фактически? Больше всего было не татарских и немецких, а польско-литовских родов.
См также дополнительная информация.
http://www.gazeta.ru/science/2008/01/14_a_2552231.shtml
А теперь опять зарубежная генетика на счет русской популяции. Dienekes Pontikos опубликовал результаты статистического анализа аутосомных SNP-маркёров 871 образцов из 60 популяций Западной Евразии. Полученное деление на кластеры выявило предсказуемую структуру, которая, тем не менее, заслуживает самого пристального внимания.Для тех, кто не хочет влезать глубоко в детали, сразу приведу структурную диаграмму с русскими, белорусами, поляками, литовцами и финнами, поставленными в один ряд с западными и южными европейцами.
Теперь доступно и подробно по делению всего исходного материала на кластеры количеством от 3 до 11. Структурная диаграмма, построенная на основе SNP-маркёров, представляет собой массив исходных данных, где каждый столбец составляет отдельный образец, либо — в данном случае — сгруппированные в популяции средние значения по образцам. Деление на кластеры проводится машинным анализом сходства фрагментов SNP-маркёров при заранее заданном количестве кластеров. Другими словами, схожие последовательности отдельных образцов объединяются в кластеры и обозначаются своими цветами.Ну и где шведы - нордманы, якобы основатели русского государства ??????? Вот синтетическая карта Европы, показывающая распределение и родство различных этнических групп:
Карта из:
http://dienekes.blogspot.ru/2010/11/clustering-of-european-y-strs.html
Четко видно что РУСЬ, ПОЛЬША, БЕЛОРУССИЯ - кровно едины !!! Русские и центрально-европейские славяне поляки - почти что одна генетическая популяция.
http://dienekes.blogspot.ru/2010/11/clustering-of-european-y-strs.html
Четко видно что РУСЬ, ПОЛЬША, БЕЛОРУССИЯ - кровно едины !!! Русские и центрально-европейские славяне поляки - почти что одна генетическая популяция.
Популяционная генетика человека включает: 1) Y - хромосома (см. Вики) - мужская линия.
2) Х - хромосома (см. Вики) - женская линия. 3) Аутосомы (см.Вики) - не половые хромосомы.
3).Аутосомами у живых организмов с хромосомным определением пола называют парные хромосомы, одинаковые у мужских и женских организмов. Иными словами, кроме половых хромосом, все остальные хромосомы у раздельнополых организмов будут являться аутосомами.
Аутосомы обозначают порядковыми номерами. Так, у человека в диплоидном наборе имеется 46 хромосом, из них — 44 аутосомы (22 пары, обозначаемые номерами с 1-го по 22-й) и одна пара половых хромосом (XX у женщин и XY у мужчин).
EUROPEAN GENETIC VARIATION - Аутосомные маркеры о русских
Карты, представляющие истоки европейского генетического разнообразия на основании аутосомных маркеров (одного из трех основных инструментов популяционной генетики, наряду с игрек-хромосомами и материнской ДНК). Составлены по исследованию: Cavalli-Sforza, L. L. (1997) Genes, Peoples and Languages. Proc Natl Acad Sci; 94:7719-7724.
Наиболее примечательны вторая, показывающая, что центр финно-угорских генов в Европе находится в северной Скандинавии, и третья, подтверждающая выводы исследований распространения игрек-хромосом о том, что прародиной индоевропейцев (ариев) являются юг и центр России.
Наиболее примечательны вторая, показывающая, что центр финно-угорских генов в Европе находится в северной Скандинавии, и третья, подтверждающая выводы исследований распространения игрек-хромосом о том, что прародиной индоевропейцев (ариев) являются юг и центр России.
Расселение неолитических земледельцев
THE FIRST component is almost superimposable to the archaeological dates of the spread of farming from the Middle East between 10,000 and 6,000 years ago.
Финно-угры
THE SECOND principal component parallels a probable spread of Uralic people and/or languages to the northeast of Europe [c. 4,000 years ago].
Арии
THE THIRD is very similar to the spread of pastoral nomads (and their successors) who domesticated the horse in the steppe towards the end of the farming expansion, and are believed by some archaeologists and linguists to have spread most Indo-European languages to Europe.
Греки
THE FOURTH is strongly reminiscent of Greek colonization in the first millennium B.C.
Баски
THE FIFTH corresponds to the progressive retreat of the boundary of the Basque language. Basques have retained, in addition to their language, believed to be descended from an original language spoken in Europe, some of their original genetic characteristics.
(Cavalli-Sforza, 1997)
Родственники русских по ДОМИНИРУЮЩЕЙ игрек хромосоме:
Лужичане, Поляки, Украинцы, Белорусы, Венгры, Чехи, Словаки, Латыши, Литовцы, Киргизы, Алтайцы, Таджики, Ишкашим, Белуджи, Пенджабцы, Пуштуны. Однотипные Хромосомы сгруппированы в гаплоидные группы, - свидельствующие о родстве народов, всего их 18. Вот например, некоторые из них.
Доминирующие мужские гаплогруппы Европы, детализация гаплогрупп русского народа из Европедии - Карта.
http://www.eupedia.com/europe/european_y-dna_haplogroups.shtml
Общепринятые Cубклады гаплогруппы R1a
Частотность гаплогруппы R1a на землях Евразии.
Изменчивость на разных уровнях популяционной иерархии.
Карта западно-евразийской гаплогруппы R1a показывает ее доминирование в
Восточной Европе и высокие частоты также в горах Алтая и Тянь-Шаня (где снижены
частоты восточно-евразийских гаплогрупп). Такая «альтернативность» гаплогруппы R1a и
восточно-евразийских гаплогрупп создает необычный кольцевой ареал R1a.
Филогенетическая сеть изменчивости этой гаплогруппы (рис. ниже ) подтверждает
существование древних кластеров STR-гаплотипов R1a в южносибирских популяциях.
Можно предполагать, что первоначально R1a была распространена по степной полосе
вплоть до Монголии. Эта зона согласуется с ареалом скифо-сарматских групп.
А это другая европейская гаплогруппа I (I1+I2), прародина которой была некогда Русская Равнина,- и её частоты в Европе и на Руси.
Обнаружение четких географических закономерностей, с одной стороны, развивает тенденции, показанные ранее . С другой стороны, основной вывод этих авторов состоял в том, что географическое соседство, а не лингвистическое родство задает структуру генофонда. Результаты исследования, О.Балановского подтверждая важность
географического фактора, указывают, что доминирует структурированность по
лингвистическому принципу. Это следует из результатов многомерного шкалирования. См.Рис.
В этот вид анализа включены те народы, для которых имелись данные по четырем или более региональным популяциям с объемом выборки не менее 70 человек.
В данном случае этнический фактор совпадает с лингвистическим, поскольку этническая принадлежность популяции определялась по языку. График показывает, что во всех случаях популяции, относящиеся к одному народу, кластеризуются совместно, образуя «этнические облака», почти не перекрывающиеся друг с другом. См.рис выше.
Два исключения касаются славянских групп. Во-первых, северные русские
популяции не присоединяются к южным, а образуют отдельный кластер. Во-вторых,
популяции поляков, южных русских и украинцев формируют не три отдельных, а одно единое «этническое облако», демонстрируя общность генофонда. Таким образом, славяноязычные популяции, занимающие половину территории Европы, оказываются генетически не различными (как должно бы быть в случае доминирования в структуре генофонда географического фактора), а сходными, в согласии с лингвистическими данными о близком родстве восточнославянских и западнославянских языков
Поскольку этнический фактор оказался столь существенным для структурирования
европейского генофонда, был проведен анализ межпопуляционной изменчивости на двух уровнях популяционной иерархии: а) внутриэтническом (различия популяций в пределах одного народа, т.е. оценка гетерогенности этнического генофонда); б) межэтническом (различия между усредненными этническими частотами гаплогрупп разных народов Европы).
Таблица показывает, что в пределах Европы наибольшая гетерогенность
характерна для финнов, хорватов, русских и итальянцев; шведы и немцы демонстрируют умеренную гетерогенность; другие народы (греки, турки, поляки, белорусы, украинцы)
характеризуются сниженной в масштабе Европы гетерогенностью. В среднем,
гетерогенность типичного европейского народа составляет 0.03 (Gst=0.029, Fst=0.027).
Таким образом, по гаплогруппам Y-хромосомы для населения Европы различия между популяциями одного этноса значительно меньше, чем различия этносов между собой (межэтнический уровень Gst =0.15, Fst =0.17). Общая величина межпопуляционных различий
по Y-хромосоме (суммарно внутриэтнический и межэтнический уровни) составляет для Европы около 0.19. Это является дополнительным подтверждением резко выраженных
различий внутри Европы по Y-хромосоме , поскольку по другим генетическим системам межпопуляционная изменчивость Европы на порядок ниже: Gst =0.02.
Балановский О.П. ИЗМЕНЧИВОСТЬ ГЕНОФОНДА В ПРОСТРАНСТВЕ И ВРЕМЕНИ:СИНТЕЗ ДАННЫХ О ГЕНОГЕОГРАФИИ МИТОХОНДРИАЛЬНОЙ ДНК И Y-ХРОМОСОМЫ.
Родственники русских по Доминируюшей икс хромосоме мтДНК.
Генетическая карта мтДНК показана на рисунке ниже, разным цветом показаны разные гены. У человека ее размер составляет 16 500 нуклеотидов.
При сравнительном изучении мтДНК у народов выявленные гаплотипы были отнесены к определенным линиям в соответствии с классификацией Макуолли. Для большинства расовых и географических групп установлена специфичность типов мтДНК (рис. ниже). Более 90% всех типов мтДНК народов Западной Евразии принадлежит к линиям Н, J, V, 1, К, Т, U, W и X, которые рассматриваются как потомки европейского генного пула, существовавшего в верхнем палеолите. Митохондриальный геном азиатских популяций включает два суперкластера М и N, которые соединяются в африканском макрокластере L3. Последний рассматривается в качестве общего предка для всех неафриканских популяций мира. Около 50% линий мтДНК коренных жителей Азии принадлежит к суперкластеру М, который подразделяется на линии С, Z, D, G, Е и др.
. Суперкластер N в Азии делится на несколько линий: А, Y, В, F
 | Рис. Медианная сеть, демонстрирующая полиморфизм мтДНК в популяциях Европы, Азии и Африки
Линии D, С, Z, G суперкластера М, линии A, Y суперкластера N и линии В, F суперкластера R характерны для азиатских популяций; линии 1, X, W суперкластера N и линии V, Н. Т, J, U, К суперкластера R - для европейских; линии LI, L2, L3, М1 и U6 принадлежат африканским народам
|
Или более наглядно все линии мтДНК
Карты распеределения этих самых "женских" гаплогрупп ДНК на территории Восточной Европы .
распределение гапплогруппы V
распределение гаплогруппы U5
распределение гапплогруппы H
Следы смешения народов можно найти в их генах. На рисунке частоты европейских и азиатских линий мтДНК для некоторых народов, населяющих Россию.
Подобное же единообразие демонстрирует и сравнение МитоДНК у русского и немецкого народов. Данные взяты из Европедии. Явно генетика у немок славянская, что позволяет сделать некоторые выводы .....
ИЗМЕНЧИВОСТЬ МИТОХОНДРИАЛЬНОЙ ДНК В ЕВРОПЕ
Изменчивость мтДНК изучена в 22 популяциях русских, украинцев и белорусов,
охватывающих все части обширного ареала восточных славян.
Частоты гаплогрупп приведены в таблице ниже. Данные представлены также на сайте www.genofond.ru .Использование этой базы данных MURKA расширяет число доступных для анализа популяций (см. табл. ).
Такое обилие данных позволило провести строгий отбор популяций по следующим критериям: а) объем выборки не менее 70 (меньшие выборки были исключены или объединены); б) надежное определение гаплогрупп (подтвержденных ПДРФ-анализом или однозначно прогнозируемых по ГВС1); в) наличие четкой географической привязки в описании выборки. Был также проведен и строгий отбор гаплогрупп: в анализ включены только гаплогруппы, средняя частота которых в Европе не менее 1% (критерий полиморфизма).
КЛАСТЕРИЗАЦИЯ ПОПУЛЯЦИЙ ПО ЧАСТОТАМ ГАПЛОГРУПП.
При анализе методом многомерного шкалирования (рис. А) четко выделились четыре кластера. Первый кластер включил только саамов, что не удивительно, учитывая их генетическое своеобразие. Второй кластер включил те популяции восточных рубежей Европы, у которых повышена частота восточно-евразийских гаплогрупп.
Третий кластер включил популяции Передней Азии и Кавказа. Все остальные популяции с
основной территории Европы (от Волги до Пиренейского полуострова) вошли в четвертый
«паневропейский» кластер, небольшие размеры которого на графике свидетельствуют о низкой межпопуляционной изменчивости. Эти результаты подтверждают гомогенность генофонда Европы , но указывают на своеобразие генофондов Приуралья и Передней Азии.
При отдельном анализе только тех популяций, которые вошли в «паневропейский»
кластер (рис. Б), обнаружилась обособленность популяций северо-востока Европы (главным образом, финно-угорских). Однако подавляющее большинство популяций по-прежнему группировалось в единый гомогенный кластер, который можно назвать «узко-европейским» (рис. Б).
европейского» кластера выявился целый ряд кластеров второго порядка. Популяции,
говорящие на языках одной лингвистической группы, расположились в генетическом
пространстве рядом друг с другом. Так, популяции германской, славянской, романской и балтской языковых групп формируют каждая отдельную группу и в генетическом
пространстве. Учитывая, что население Шотландии лишь исторически недавно сменило язык, их генетическое сходство с ирландцами обрисовывает общность митохондриального генофонда и кельтской группы (рис. В).
В обнаруженной закономерности генофонда Европы (популяции одной языковой
группы характеризуются сходными митохондриальными генофондами) отмечаются два исключения: румыны и аромуны (лингвистически принадлежат к романской группе, но генетически обособлены), а также эстонцы и венгры (лингвистически финно-угорская группа, но генетически примыкают к «славянскому» кластеру).
РОЛЬ ЛИНГВИСТИЧЕСКОГО И ГЕОГРАФИЧЕСКОГО ФАКТОРОВ.
Значимость географического и лингвистического факторов оценивалась во многих работах . Впервые обнаруженная значимость лингвистической принадлежности популяций для структурирования митохондриального генофонда Европы подтверждена и другим методом: проведен анализ связи генетических, географических и лингвистических расстояний между популяциями Европы (тест Мантеля, табл. ниже). Выявлен примерно одинаковый уровень корреляции генетических расстояний и с географическими, и с лингвистическими расстояниями.
ИЗМЕНЧИВОСТЬ В ПРЕДЕЛАХ ЛИНГВИСТИЧЕСКИХ ГРУПП.
Для уточнения выявленной связи генетических кластеров популяций с лингвистическими группами была рассчитана межпопуляционная изменчивость (гетерогенность) в пределах каждой из лингвистических групп Европы (табл. , рис. ).
ГАПЛОТИПИЧЕСКОЕ РАЗНООБРАЗИЕ мтДНК.
Все выше описанные виды анализа
проведены по частотам гаплогрупп. Для увеличения разрешающей способности проведен также анализ и по отдельным гаплотипам. Уровень гаплотипического разнообразия рассчитан для 51 восточноевропейских популяций как по собственным данным, так и по созданной базе данных . Столь представительный анализ гаплотипического разнообразия мтДНК выполнен впервые в мире. Картографирование этого показателя демонстрирует закономерное убывание к северу: от 0.99 у популяций Причерноморья до 0.95 и ниже на берегах Северного Ледовитого океана (рис. ниже).
Можно предположить причинно-следственную цепь взаимосвязанных изменений от юга к северу сразу нескольких параметров: географической широты (возрастает),
продуктивности среды (убывает), плотности населения (убывает), эффективного размера
популяции (убывает), изоляции субпопуляций (возрастает), дрейфа генов (возрастает),
гаплотипического разнообразия мтДНК (убывает).
Согласно теории популяционной генетики интенсивный дрейф генов должен приводить
к возрастанию случайного инбридинга, а, следовательно, и генетического груза популяции. И действительно, выявлена достоверная корреляция (r=0.4) между гаплотипическим разнообразием мтДНК и отягощенностью популяций аутосомно-рецессивной наследственной патологией . Это подтверждает роль дрейфа генов как ключевого фактора в формировании северных генофондов.
ОСОБЕННОСТИ ГЕНОФОНДА ВОСТОЧНЫХ СЛАВЯН
Структура генофонда изучена как на уровне Европы и Евразии в целом, так и на более
низких иерархических уровнях. В качестве такого модельного объекта выбраны восточные
славяне (наиболее многочисленная группа населения России и сопредельных стран
Европы), а среди восточных славян особое внимание уделено русскому этносу.
МИТОХОНДРИАЛЬНЫЙ ГЕНОФОНД ВОСТОЧНЫХ СЛАВЯН.
Исследованию изменчивости мтДНК у восточных славян в их историческом ареале посвящен ряд работ , различавшихся подходами к формированию выборок. Поскольку для близкородственных генофондов этот вопрос особенно важен, то в анализ включены главным образом собственные данные О.Балановского. Результаты анализа частот гаплогрупп у 22 восточнославянских популяций (N=2 396 образцов) и окружающих их неславянских народов представлены на рис. ниже.
Во избежание перегрузки графика, на нем оставлены лишь популяции (от Балканского полуострова до Урала), имеющие непосредственное отношение к этногенезу восточной ветви славян. Выявленная структура генофонда сводится к трем кластерам. В «приуральский» кластер вошли восточные финно-угорские (марийцы, коми) и тюркские народы (татары, чуваши). В «восточно-европейский» кластер вошли западные и восточные славяне, балты, а также балканские народы. В «северо-европейский» кластер вошли западные финно-угорские народы и северные русские популяции.
Таким образом, выявляется генетически гомогенная балто-славянская общность, к
которой тяготеют также и балканские популяции («восточно-европейский» кластер). От этой общности отличаются финно-угорские и тюркские народы Восточной Европы, генофонд которых, в свою очередь, подразделяется на западную и восточную части (кластеры 1 и 3).
Такая картина могла сформироваться в ходе славянской колонизации Восточно-Европейской равнины : двигающееся с запада славянское население несло с собой «восточно-европейский» митохондриальный генофонд, который замещал ранее существовавший на этих территориях автохтонный «приуральский» генофонд, разделяя оставшиеся не ассимилированными группы на западную и восточную части.
ГЕНОФОНД ВОСТОЧНЫХ СЛАВЯН ПО Y-ХРОМОСОМЕ.
Анализ Y-хромосомы обрисовал в целом аналогичную картину: восточнославянские популяции оказываются сходными между собой и с западнославянскими популяциями, при этом прослеживается своеобразие северных русских популяций . Различия в результатах, полученных по мтДНК и Y-хромосоме, невелики и связаны
главным образом с тем, что мтДНК в целом более гомогенна в Европе и поэтому в меньшей
степени, чем Y-хромосома, выявляет различия между генофондами. Например, по мтДНК
южные славяне присоединяются к остальной славянской общности, хотя в генетическом
пространстве располагаются на ее окраине (рис. выше). Но результаты по Y-хромосоме
(разрешающая способность которой выше) уточняют картину: четко видно, что южные
славяне отделены от восточных и западных славян . Ряд закономерностей выявлен для генофондов украинцев и белорусов. Литературные данные об этих популяциях получены по трудно сопоставимым панелям SNP маркеров, поэтому мы опирались главным образом на собственные данные. Для украинцев показано сходство с более северными группами (Польши, Белоруссии и юга России) и некоторое своеобразие западных украинцев. Обнаружена гомогенность генофонда белорусов, сквозь которую все же пробиваются слабые различия между северными и южными группами .
СТРУКТУРА РУССКОГО ГЕНОФОНДА.
Более высокая разрешающая способность Y-хромосомы по сравнению с мтДНК обнаруживается и при анализе русского генофонда. По мтДНК его структура выявляется лишь на уровне тенденций: в изменении частот некоторых гаплогрупп с юга на север, в отличии окраинных популяций казаков и поморов (Балановский и др., 2009). Близкие результаты получены и другими авторами . При анализе же Y-хромосомы изменчивость большинства гаплогрупп близка к клинальной, что подтверждено анализом автокорреляций . Информативность Y-хромосомы проявляется и при многомерном анализе: центральные и южные популяции оказываются генетически сходными, а северные популяции - своеобразными (рис. ниже).
Кардинальные различия северных и южных русских проявляются и при анализе карт
главных компонент изменчивости различных признаков: гаплогрупп Y-хромосомы и
классических маркеров , фамилий ,
антропологических признаков . Взаимное подтверждение результатов по четырем системам признаков (принцип «полисистемного подхода») не
оставляет сомнений в реальности обнаруженной закономерности широтной изменчивости русского генофонда.
Первая причина высокой межпопуляционной изменчивости на Русском Севере - сниженная плотность населения и меньший эффективный размер популяций . Поэтому дрейф генов в
северных русских популяциях действует сильнее. Тем самым снова обнаруживается
параллелизм с изменчивостью мтДНК (с описанным выше широтным трендом
гаплотипического разнообразия и связанных с ним параметров). Но отличия северных
русских популяций вызваны не только действием дрейфа генов: в этом случае северные русские группы разместились бы случайным образом вокруг южных, без систематического сдвига в одну сторону. Поэтому второй причиной могут быть особенности происхождения северных русских. Это побудило нас провести специальное исследование генофонда Русского Севера .
ПРОИСХОЖДЕНИЕ СЕВЕРНЫХ РУССКИХ ПОПУЛЯЦИЙ.
Диаграмма-график родства русского народа со скандинавами и конкретно со шведами.
http://www.ncbi.nlm.nih.gov/pmc/articles/PMC2730349/?tool=pubmed
Сходство Русского Севера с географически отдаленными балтами более выражено, чем с финно-уграми. Этот пестрый в лингвистическом отношении (славяне, балты, финно-угры, германцы), но географически единый (от Балтики до Печоры) континуум популяций мог сформироваться еще до выделения балто-славянских и финских лингвистических групп. По митохондриальной ДНК Русский Север обнаруживает близость с еще более отдаленными популяциями Европы (рис. ниже).
Русский Север оказался генетически близким к
норвежцам, немцам, полякам, литовцам и т.д., и при этом генетически отличным от
финноязычных популяций. Анализ аутосомного маркера CCR5del32, изменчивость которого активно изучалась многими российскими коллективами дал сходные результаты .
В целом, обнаруженное сходство северных русских с популяциями Северной и
Центральной Европы позволяет выдвинуть гипотезу о сохранении на этих территориях
древнего генофонда Европы, восходящего, возможно, еще к эпохе мезолита. Этот
палеоевропейский генофонд унаследован как северными древнерусскими популяциями, так
и частью финноязычных племен, расселившихся здесь, а также балтоязычными и германоязычными популяциями.
http://www.med-gen.ru/ar/ar_Balanovsky.pdf
Распределение частот гаплогрупп мтДНК у русских согласно http://www.eupedia.com/europe/european_mtdna_haplogroups_frequency.shtml
Multiple origins of Russian mtDNA
First PC of mtDNA variation on the left. From the paper:
The genetic distances from the Russians to the Europeanlanguage groups indicate that the gene pool of present-day Russians bears the influence of Slavic, Baltic,Finno-Ugric and, to a lesser extent, Germanic groups, aswell as Iranian and Turkic groups....
The results of this study strongly suggest that the impact of the pre-Slavic (Finno-Ugric) population on the East European Plain is the most important factor for the northward and southward differentiation of the present-day Russian gene pool. This explanation supports the view proposing the genetic influence of Finno-Ugrians on the formation of the northern regions of Russia, which was inferred from mtDNA marker studies of some Russian populations (Grzybowski et al., 2007) and Y-chromosome analysis (Balanovsky et al., 2008).
Being quite distant from the Finno-Ugric group, the Southern Russians consequently differ from the Northern Russians in their closeness to the Germanic group. This difference indicates that the Germanic people played a significant role in the development of the southern, but not the northern segment of the Russian gene pool. In general, the Germanic influence on the formation of the Russians is not as obvious as the impact of the Slavic, Baltic, and Finno-Ugric people. However, strong interactions between the Germanic and Slavic tribes have been found in archeological materials dating from the mid-first millennium B.C. to the early first millennium A.D. These interactions were the strongest on the northern coast of the Black Sea, in the area of the multiethnic Chernyakhov archeological culture (second to fifth centuries A.D.). In the second half of the first millennium A.D., the descendants of this culture colonized the southern regions of the historical Russian area (Sedov, 1994, 1995). However, there is no evidence in the historical literature of the interaction between the Germanic tribes and the Slavs (and later, the Russians) after the Slavic colonization of the East European Plain. Therefore, the Germanic influence could not have occurred after the early part of the first millennium A.D., which was before the eastward Slavic migration (Sedov, 1994, 1995). Apparently, the impact of the Germanic people on the Chernyakhov Slavs affected the gene pool of modern Southern Russians, consequently differentiating them from the Northern Russians (Fig. 6).Am J Phys Anthropol DOI: 10.1002/ajpa.21649
Russian ethnic history inferred from mitochondrial DNA diversity
Irina Morozova et al.
With the aim of gaining insight into the genetic history of the Russians, we have studied mitochondrial DNA diversity among a number of modern Russian populations. Polymorphisms in mtDNA markers (HVS-I and restriction sites of the coding region) of populations from 14 regions within present-day European Russia were investigated. Based on analysis of the mitochondrial gene pool geographic structure, we have identified three different elements in it and a vast “intermediate” zone between them. The analysis of the genetic distances from these elements to the European ethnic groups revealed the main causes of the Russian mitochondrial gene pool differentiation. The investigation of this pattern in historic perspective showed that the structure of the mitochondrial gene pool of the present-day Russians largely conforms to the tribal structure of the medieval Slavs who laid the foundation of modern Russians.Our results indicate that the formation of the genetic diversity currently observed among Russians can be traced to the second half of the first millennium A.D., the time of the colonization of the East European Plain by the Slavic tribes. Patterns of diversity are explained by both the impact of the native population of the East European Plain and by genetic differences among the early Slavs.
Link
Родственники русских по Аутосомам - не половым хромосомам.
Полиморфизм классических генетических маркеров
По данным о классических генных маркерах , как и по диаллельным ДНК маркерам, Евразийский генофонд подразделяется на «западный» и «восточный» миры (рис. ниже ). Все четыре Кавказских региона, как и при анализе диаллельных ДНК маркеров, образуют очень компактный самостоятельный кластер, близкий к народам Юго-Западной Азии. Во вторую компактную группу регионов, генетически близкую к Кавказу, вошли популяции Европы – Южной, Западной. Третью группу регионов образуют генофонды Урала - Приуралье и Южный Урал. К ним генетически относительно близки народы Зауралья и Центральной Азии, но они уже принадлежат к суперкластеру «Восточная Евразия».
Рис. Положение региональных генофондов Евразии по девяти классическим генным маркерам (ABO, RH, ACP1, ESD, HP, GC, TF, GLO1, С’3)
Примечание: дендрограмма построена методом Уорда
Из схемы видно что Восточная Европа более удалена генетически от Кавказа, Центральной и Южной Азии чем Западная... Западноевропейцы более БЛИЗКИ кавказоидам и среднеазиатам чем славяне!!!
Из схемы видно что Восточная Европа более удалена генетически от Кавказа, Центральной и Южной Азии чем Западная... Западноевропейцы более БЛИЗКИ кавказоидам и среднеазиатам чем славяне!!!
Диаллельный ДНК полиморфизм
Результаты, полученные разными методами анализа (кластерный, факторный, многомерное шкалирование), в целом согласуются, т.е. выявленное положение этносов в генетическом пространстве не зависит от метода анализа.
Рисунок ниже убедительно демонстрирует высокую согласованность генетического и лингвистического сходства. В «славянский» кластер вместе с кубанскими казаками вошли русские и украинцы. «Абхазо-адыгский» кластер объединил все народы, относящиеся к абхазо-адыгской группе северокавказской языковой семьи – абазин, абхазов, адыгейцев, кабардинцев, черкесов. В «нахско-дагестанский» кластер вошли чеченцы и народы Дагестана, относящиеся к нахско-дагестанский группе северокавказской языковой семьи.
Отличия от лингвистической классификации обнаруживают лишь тюркоязычные народы Кавказа, однако их положение на генетическом древе полностью согласуется с особенностью их этногенеза. Показательно, что тюркские народы высокогорных районов Кавказа (балкарцы и карачаевцы) вошли в «нахско-дагестанский» генетический кластер. Известно, что они относительно недавно восприняли тюркские языки – ранее они были ираноязычны, причем предполагается, что до этого значительная часть их предков говорила на языках северокавказской языковой семьи. К этому же кластеру присоединилось и тюркоязычное население предгорий Дагестана – кумыки, однако они генетически далеки от единого массива высокогорных популяций (рис. ниже).
Этносы «степных тюрок» (ногайцы и караногайцы) образовали свой собственный генетически резко отличающийся кластер. Близость тюркоязычных горцев к горцам нахско-дагестанской группы, а не к «степным тюркам», соответствует их этногенезу.
Удивительно, что кубанские казаки генетически оказались в ином кластере, чем адыги. Казачье население на Северном Кавказе сформировалось в середине XIX века и в течение ряда поколений казаки проживают в тесном соседстве с адыгами. Однако установленный нами факт значительных генетических различий между генофондами казаков и адыгов позволяет утверждать, что поток генов между их генофондами невелик.
Рис. Положение генофонда славянских народов относительно других народов Кавказа по семи диаллельным аутосомным ДНК маркерам (АСЕ, PV92, TPA25, ApoA1, А25, В65, CCR5del32)
Примечание: дендрограмма построена методом Уорда
Полиморфизм мультиаллельных ДНК маркеров
Несмотря на то, что по мультиаллельным ДНК маркерам изучена иная панель народов, паттерн изменчивости русского генофонда полностью согласуется с полученным по диаллельным ДНК маркерам. Мы вновь видим (рис. ниже), что популяция кубанских казаков вошла в общий кластер с русскими и украинцами, а не адыгейцами, рядом с которыми кубанские казаки проживают в течение шести поколений. Абхазо-адыгские народы также образовали свой собственный генетический кластер. Степные тюрки - ногайцы и караногайцы – образовали общий кластер, включающий также башкир Южного Урала.

Рис. Положение генофонда славянских народов среди других соседних народов по
мультиаллельным аутосомным ДНК маркерам (D1S80,
ApoB, DM)
Примечания: диаграмма
двумерного шкалирования; число итераций–34, величина стресса Sо=0,10; коэффициент
алиенации Ко=0,13; кривая Шепарда удовлетворительна
Рис. Положение генофонда славянских народов среди других соседних народов по девяти классическим генным
маркерам (ABO, RH, ACP1, ESD,
HP, GC, TF, GLO1, С’3)
Примечания: диаграмма двумерного шкалирования; число итераций–34, величина стресса Sо=0,13; коэффициент алиенации Ко=0,158; кривая Шепарда
удовлетворительна.
Смотриhttp://www.med-gen.ru/ar/ar_PocheshovaEA.doc
Биологические, археологические и культурологические доказательства палеоазиатского происхождения северных монголоидов, европеоидов и американских индейцев
Ученые России обнаружили европеоидные частоты генов белков и ферментов крови (А.Ф.Назарова, М.Г.Кузнецова. Генетическая структура популяций алтайцев. Доклады РАН, 1993, 333, 3, 405-409.) в семи популяциях алтайцев. Вычисление генетических расстояний 11 человеческих популяций Европы, Азии и Америки, а именно саамов (лопарей), ненцев, нганасан, эвенков, якутов, монголов, алтайцев, русских, финнов, немцев и американских индейцев по частотам 28 аллелей белков, ферментов и групп крови показало определенную близость по наследственным признакам европеоидов и северных монголоидов (Табл.ниже).
К русским ближе всего оказались из славянских народов поляки, как и при вычислении генетических расстояний по меньшему числу генетических локусов .В одном большом кластере с русскими оказались иранцы, коми, чуваши, удмурты, ненцы,и субкластер осетин и азербайджанцев.  1. Русские 2. Поляки 3. Иранцы 4. Коми 5. Чуваши 6. Удмурты 7. Ненцы 8. Осетины 9. Арабы 10. Итальянцы 11. Немцы 12. Финны 13. Алтайцы 14. Эвенки 15. Якуты 16. Монголы 17. Индийцы 18. Украинцы 19. Белорусы 20. Эстонцы 21. Шотландцы 22. Сербы 23. Хорваты 24. Болгары 25. Чехи 26. Венгры 27. Марийцы 28. Шведы 29. Манси 30. Индейцы Америки 31. Татары 32. Евреи Ближнего Востока 33. Негры Африки 34. Греки 35. Лопари
1. Русские 2. Поляки 3. Иранцы 4. Коми 5. Чуваши 6. Удмурты 7. Ненцы 8. Осетины 9. Арабы 10. Итальянцы 11. Немцы 12. Финны 13. Алтайцы 14. Эвенки 15. Якуты 16. Монголы 17. Индийцы 18. Украинцы 19. Белорусы 20. Эстонцы 21. Шотландцы 22. Сербы 23. Хорваты 24. Болгары 25. Чехи 26. Венгры 27. Марийцы 28. Шведы 29. Манси 30. Индейцы Америки 31. Татары 32. Евреи Ближнего Востока 33. Негры Африки 34. Греки 35. Лопари
http://www.trinitas.ru/rus/doc/0016/001b/00161386.htmhttp://www.trinitas.ru/rus/doc/0211/002a/1087-naz.pdf
Ради интереса: Российский антрополог Т.В Томашевич обнаружила градиент распределения частот надглазничных каналов черепа человека (Табл.ниже)
Таблица .
Распределение частот надглазничных каналов черепа человека
В надглазничных каналах проходят надглазничные и надблоковые артерий и вен — ветвей глазничной артерии. В исключительных случаях в них проходят надглазничная и надблоковая ветви лобного нерва. Как видно из данных Табл. , у саамов, американских индейцев, а также северных монголоидов - якутов, юкагиров, эвенков, чукчей, эскимосов наиболее высокая частота встречаемости надглазничных каналов, что возможно, вызвано их палеоазиатским происхождением. У кетов, юкагиров, русских, манси, осетинов, армян частота надглазничных каналов очень близка и варьирует в пределах 30-38 %, это может быть подтверждением их общего азиатского происхождения. У монголоидов Восточной Азии – японцев, корейцев, китайцев, а также у монголов частота надглазничных каналов 43-48%. Интересно, что у древних египтян, этрусков, древних славян (полян киевских) и индийцев Уттар Прадеш частота каналов практически одинакова и составляет 25-28%, чуть выше она у словен новгород-ских; это также может быть связано, с происхождением этих народов из древней палеоазиатской популяции.
Итак, родственники русских по всем трём показателям (хромосомы половые и неполовые), - поляки. Русские и Поляки прямые потомки легендарных вандалов, об этом сказано даже в Википедии.
http://ru.wikipedia.org/wiki/%D0%92%D0%B0%D0%BD%D0%B4%D0%B0%D0%BB%D1%8B
Рюрик происходил из севера Германии, из Пруссии, из народов Ругов или Вандалов. На тот момент, 9 век вряд ли вандалы с ругами уже отличались. Т.е он вандал, скорей всего из города Любека с юга Балтики. Вот карта той славянской Германии
Север Германии:
А вот современная карта от Гугла:
Если посмотреть на карту Германии, то в ее северной части можно заметить много названий славянского корня. Городки: Рерик, Руссов, Руггов...С точки зрения генетики, Русские находятся между немцами и поляками, и приходяться как бы на некогда славянские земли Шлезвиг-Гольштейн (земля вандалов силингов) и Верхняя Померания (земля поморов мореплателей).
Или
Эти схемы указывают на родство русских с Северной Германией и со Скандинавией; (Опять Померания и Шлезвиг-Гольштейн, т.е славянская Силезия )
http://eurogenes.blogspot.ru/2011/02/swedish-genetic-substructure-something.html
http://eurogenes.blogspot.com/2011/05/german-sorbs-genetically-closer-to.html
Популяционная генетика подтверждает полное родство русских с народами южной Балтики, с поляками и лужичанами,- менее выраженное родство с эстонцами, латышами, литовцами. См:
http://svpressa.ru/society/article/60385/
http://www.dzd.ee/449810/analiz-dnk-u-latyshej-litovcev-jestoncev-i-russkih-obwie-geny/
Как уже было продемонстрировано подтверждает только частичное родство с народами северной Балтики: норвежцами по женской линии!.
Тесты:
Аутосомно-генетических расстояний (Fst) на основе SNPs (2009) , т.е на основе Однонуклеотидного полиморфизма
Генетические расстояния между популяциями часто измеряется индексом фиксации (Fst), основанным на генетических данных полиморфизма, таких как однонуклеотидный полиморфизм (SNP) или микросателлиты. Fst является частным случаем F-статистики, концепция, разработанная в 1920-х годах Сьюэллом Райтом.
Fst это просто соотношение случайно выбранных аллелей в пределах одной группы населения по сравнению с тем, что установлено на все население. Это часто выражается как доля генетического разнообразия за счет различия частоты аллелей между популяциями. Диапазон значений от 0 до 1. Нулевое значение означает, что две популяции взаимозаменяемы, что они свободно скрещиваются. Значение 1 будет означать, что две популяции полностью разделены. Чем больше Fst значение, тем большее генетическое расстояние. По существу, низкие значения Fst предпологают, что большинство генетических вариаций отдельных лиц находится на уровне в пределах одной группы населения (~ 85%), в то время как принадлежащие к различным группам населения находятся в пределах той же «расы» / континента, и даже при различным расовых / континентальных группах, они представлены в гораздо меньших степенях вариации (3-8%, 6-11%, соответственно).
Fst это просто соотношение случайно выбранных аллелей в пределах одной группы населения по сравнению с тем, что установлено на все население. Это часто выражается как доля генетического разнообразия за счет различия частоты аллелей между популяциями. Диапазон значений от 0 до 1. Нулевое значение означает, что две популяции взаимозаменяемы, что они свободно скрещиваются. Значение 1 будет означать, что две популяции полностью разделены. Чем больше Fst значение, тем большее генетическое расстояние. По существу, низкие значения Fst предпологают, что большинство генетических вариаций отдельных лиц находится на уровне в пределах одной группы населения (~ 85%), в то время как принадлежащие к различным группам населения находятся в пределах той же «расы» / континента, и даже при различным расовых / континентальных группах, они представлены в гораздо меньших степенях вариации (3-8%, 6-11%, соответственно).
http://en.wikipedia.org/wiki/Genetic_history_of_Europe
Исследования Калифорнийского Университета по общеевропейской генетике:
Исследования Калифорнийского Университета по общеевропейской генетике:
Для получения подобного результата потребовалось задействовать в тестировании 3192 человека из различных уголков Европы, добровольно согласившихся на ДНК-секвенирование. В ходе исследования была составлена генетическая карта Европы.
Ещё карта Джона Новембера
Novembre's team's map, showing predicted locations for each of 1,387 individuals based on their DNA. Definitions: AL, Albania; AT, Austria; BA, Bosnia-Herzegovina; BE, Belgium; BG, Bulgaria; CH-I, CH-F, and CH-G denote Swiss individuals who speak Italian, French, or German respectively; CY, Cyprus; CZ, Czech Republic; DE, Germany; DK, Denmark; ES, Spain; FI, Finland; FR, France; GB, United Kingdom; GR, Greece; HR, Croatia; HU, Hungary; IE, Ireland; IT, Italy; KS, Kosovo; LV, Latvia; MK, Macedonia; NO, Norway; NL, Netherlands; PL, Poland; PT, Portugal; RO, Romania; RS, Serbia and Montenegro; RU, Russia, Sct, Scotland; SE, Sweden; SI, Slovenia; SK, Slovakia; TR, Turkey; UA, Ukraine; YG, Yugoslavia. (Image: Nature). Можно взглянуть на карту в большем формате - вот здесь.
Важно: Карта кликабельная! А это источник!
http://www.newscientist.com/article/dn14
Не обошлось и без странностей. Так, например, россияне разместились на этой карте среди чехов и поляков, оказавшись куда «ближе к Европе», нежели украинцы.
http://www.gazeta.ru/science/2008/09/01_a_2827658.shtml
Тем не менее, пользуясь только этой картой, родину 90% человек можно указать с точностью до 700 км, а 50% — до 300 км.
Вместе с тем метод главных компонент не смог разделить европейцев на отдельные генетические кластеры. Другими словами, генетическое разнообразие жителей Европы настолько мало, что даже полумиллиона мононуклеотидных полиморфизмов недостаточно, чтобы разделить всего три тысячи человек на маленькие подгруппы.
" Русские= Местные Роксаланы + Пришлые Вандалы" (С поселений рек Дуная и Травны). Поэтому генетика русских тесно связана как с Центральной Европой так и с Восточной. Вот генетическая однородность русских славян с прибалтами и скандинавами
http://dienekes.blogspot.com/2009/02/auton-et-al-on-global-distribution-of.html
Русские, Белорусские Славяне, Прибалты и скандинавы наиболее однообразны, после британцев. Генетик и антрополог Dienekes подвёл итоги проекта по обработке SNP-материала и опубликовал дополнение, которого не хватало в недавнем кластером анализе, — наглядные диаграммы главных компонент: от первой до пятой.
Как видно, первая главная компонента отделяет Западную Евразию от Индии, вторая задаёт клинальный градиент между северными европейцами и средиземноморцами, третья выделяет из массива Западной Евразии североафриканское влияние, четвёртая — ашкеназов, пятая — финнов.Обращает на себя внимание плотная группировка русских со скандинавами, скандинавов — с немцами и ирландцами, половины немцев — с жителями Балкан, и относительная близость испанцев к северным европейцам. Показательно также, что по второму компоненту индусы находятся строго на уровне северных европейцев, не смещаясь по средиземноморской оси в сторону турок, армян и прочих греков.
Русский генофонд - русские Севера
Новое исследование вносит ясность в вызывающий много спекуляций вопрос о происхождении севернорусской популяции. Вопреки расхожему мнению, генетическое влияние финно-угров на северных русских незначительно; в действительности они являются смесью палеоевропейского субстрата со славянскими колонистами.
По данным о полиморфизме Y хромосомы (отражающей наиболее дифференцированные линии наследования) все популяции Архангельской области входят в обширный «северный» кластер вместе с популяциями вологодских русских, балтов (латышей и литовцев), финноязычных народов (коми, финнов, эстонцев и сборной группы карелов, вепсов и ижорцев), а также германоязычных шведов. При этом сходство Русского севера с географически удаленными балтами более выражено, чем с финно-уграми. В целом, генетические сходство столь пестрого в лингвистическом отношении (славяне, балты, финно-угры, германцы), но единого в географическом пространстве (от Балтики до Печоры) континуума популяций может указывать на сохранение на этой территории палеоевропейского генофонда, предшествовавшего разделению балто-славянских и финских лингвистических общностей.По митохондриальной ДНК (отражающей материнские линии наследования) Русский Север обнаруживает близость с еще более обширным массивом народов Северной Европы. Генетически наиболее близки оказались к Русскому Северу оказались норвежцы и немцы, в кластер вошли также австрийцы, швейцарцы, поляки, боснийцы, литовцы, ирландцы, шотландцы. Близкий кластер образован остальными русскими популяциями, шведами, эстонцами, латышами, украинцами , белорусами, чехами, словаками, венграми. При этом основной массив финноязычных популяций – финны, карелы, коми, мордва и мари – максимально удален и от всего европейского массива, и от Русского Севера, что отрицает предполагавшийся ранее мощный вклад финно-угорского пласта в формирование населения Русского севера.Данные по наиболее изученному и высоко полиморфному в Европе аутосомному маркеру ССR5del32 подтверждают этот вывод: область регулярно встречаемых частот ССR5del32 охватывает генофонд населения северной половины Европы, указывая на существование мощного общего генетического пласта, включающего в себя и Русский Север. Причем по числу народов с высокой частотой ССR5del32 финноязычные народы уступают и германоязычным, и славянским.
В целом, обнаруженное сходство генофонда Русского Севера с генофондами Центральной Европы и севера Восточной Европы позволяет выдвинуть гипотезу о сохранении на этих территориях древнего генофонда Европы, восходящего, возможно, еще к эпохе мезолита. Этот палеоевропейский генофонд был унаследован как северными древнерусскими популяциями, таки частью финноязычных племен, расселившихся здесь в более раннее время. Полученные данные позволяют предположить, что палеоевропейское население севера Восточной Европы, на основе которого сформировалось также балтское и германоязычное население современной Европы, длительное время сохранялось в регионах, прилегающих к Белому морю, и в эпоху раннего средневековья испытало мощное влияние северной волны древних славян. Поэтому ответ, который молекулярная генетика в настоящее время может дать на вопрос, вынесенный в заголовок статьи, можно сформулировать таким образом: «генофонд Русского Севера = палеоевропейцы + северная волна славянской колонизации».
Материалы и методы
Для создания генетического портрета населения Архангельской области использована вся доступная нам информация о полиморфизме четырех систем признаков - Y хромосомы, митохондриальной ДНК (мтДНК), аутосомных ДНК маркеров и квазигенетических маркеров - фамилий (табл. 1, рис. 1). Использовались как архивные и опубликованные данные лаборатории популяционной генетики человека МГНЦ РАМН, так и опубликованные данные других авторов. В контексте данной работы термины «генофонд населения Архангельской области» и «генофонд Русского Севера» использованы как синонимы. ДНК коллекции лаборатории популяционной генетики человека Медико-генетического научного центра РАМН (далее сокращенно обозначается ЛПГЧ), представляющие основной исторический ареал русского народа, генотипировались О.П. Балановским по мтДНК и частично по Y хромосоме в Эстонском биоцентре РАМН (рук. Р. Виллемс), остальная часть данных по Y хромосоме и данные по аутосомным ДНК маркерам получены в ЛПГЧ (табл. 1).
Для сравнительного анализа с другими русскими популяциями и народами Европы использована информация из созданных нами баз данных по мтДНК и Y хромосоме (табл. 2). Эти две системы маркеров являются в настоящее время наиболее изучаемыми: ежегодно в мире публикуются десятки популяционно-генетических работ, посвященных анализу мтДНК и Y хромосомы в популяциях мира. Коллектив ЛПГЧ в течение ряда лет ведет большую работу по сбору и систематизации этой информации в рамках специальных баз данных (БД). Так, БД «МУРКА» (база данных по митохондриальной ДНК и интегрированное программное обеспечение, основные составители: В.В. Запорожченко, О.П. Балановский, А.С. П-шеничнов, Е.В. Балановская) в 2004 году содержала информацию о 33 000 образцах мтДНК; в 2007 году - о 62 000; в 2008 году - о 95 000; в 2010 году - о 135 000, на текущий момент - о 170 000 образцах.
<☡ href="http://aquilaaquilonis.livejournal.com/2Ⓒ2902.html" styl␥="color: rgb(0, 0, 0) !important; margin: 0px;"⚮http://aquilaaq␅ilonis.livejournal.com/282902.html

Американские генетики из университета Пенсильвании (Penn State) в стремлении найти гены, связанные с раком, начали исследование аквариумных рыбок данио. В итоге они изолировали ген, который отвечает за цвет человеческой кожи.
Учёные пытались идентифицировать этот ген в течение многих лет, но в результате предыдущих исследований был обнаружен не один, а больше 100 различных генов, вовлечённых в производство меланина — пигмента, ответственного за цвет кожи.
Генетик Кейт Чэн (Keith C. Cheng) вместе с коллегами предположил, что золотая данио — более светлая версия её «дикого» сородича — имеет генетическую мутацию, из-за которой страдает белок, необходимый для производства меланина.
Оказалось, что простое добавление нормального белка сделало окрас рыбки более тёмным. Когда же учёные использовали человеческий ген SLC24A5, ответственный за производство того же белка у людей, эмбрионы рыбок также стали темнее.
Используя базу данных по геному человека (HapMap), исследователи узнали, что SLC24A5 имеет всего две вариации: у европейцев присутствует версия гена с одним типом аминокислоты (треонин), а у всех остальных — с другим (аланин).
Исследование генетики 308 человек смешанного европейского и африканского происхождения показало, что люди с треониновым вариантом SLC24A5 имеют наиболее светлую кожу, а с аланиновым типом — самую тёмную. Цвет кожи людей с обеими версиями гена находится где-то посередине между двумя «крайностями».
Группа Чэна заключила, что от изменений версий SLC24A5 зависит 25-38-процентное различие по цвету кожи между европейцами и африканцами.
Пигментация кожи у русских:
Ген цвета кожи человека
Открытие этого гена, однако, не отвечает на вопрос, почему у европейцев более светлая кожа. Хотя учёные полагают, что всё дело в повышении производства витамина Д в лишённых солнцем широтах (иллюстрация с сайта sciam.com).
Американские генетики из университета Пенсильвании (Penn State) в стремлении найти гены, связанные с раком, начали исследование аквариумных рыбок данио. В итоге они изолировали ген, который отвечает за цвет человеческой кожи.
Учёные пытались идентифицировать этот ген в течение многих лет, но в результате предыдущих исследований был обнаружен не один, а больше 100 различных генов, вовлечённых в производство меланина — пигмента, ответственного за цвет кожи.
Генетик Кейт Чэн (Keith C. Cheng) вместе с коллегами предположил, что золотая данио — более светлая версия её «дикого» сородича — имеет генетическую мутацию, из-за которой страдает белок, необходимый для производства меланина.
Оказалось, что простое добавление нормального белка сделало окрас рыбки более тёмным. Когда же учёные использовали человеческий ген SLC24A5, ответственный за производство того же белка у людей, эмбрионы рыбок также стали темнее.
Используя базу данных по геному человека (HapMap), исследователи узнали, что SLC24A5 имеет всего две вариации: у европейцев присутствует версия гена с одним типом аминокислоты (треонин), а у всех остальных — с другим (аланин).
Исследование генетики 308 человек смешанного европейского и африканского происхождения показало, что люди с треониновым вариантом SLC24A5 имеют наиболее светлую кожу, а с аланиновым типом — самую тёмную. Цвет кожи людей с обеими версиями гена находится где-то посередине между двумя «крайностями».
Группа Чэна заключила, что от изменений версий SLC24A5 зависит 25-38-процентное различие по цвету кожи между европейцами и африканцами.
Пигментационная генетика европейцев
Новое генетическое исследование аллелей, отвечающих за светлый цвет волос и глаз у европейцев, лишний раз свидетельствует, что русские входят в число самых белых из белых людей.


Источник: Pigmentation genetics of Europeans.
Источник: Pigmentation genetics of Europeans.
Пигментация кожи у русских:
So who's the most European of us all?
Lithuanians 77%
Finns 74%
Belorussians 70%
Swedes 69%
Norwegians 68%
Vologda Russians 68%
Russians 68%
Poles 68%
Erzya 66%
Ukrainians 66%
Moksha 66%
Orcadians 63%
HapMap Utah Americans (CEU) 63%
Irish 63%
British 62%
Finns 74%
Belorussians 70%
Swedes 69%
Norwegians 68%
Vologda Russians 68%
Russians 68%
Poles 68%
Erzya 66%
Ukrainians 66%
Moksha 66%
Orcadians 63%
HapMap Utah Americans (CEU) 63%
Irish 63%
British 62%
У вологодцев насчитали самый большой % северных генов, у русских из центральных областей он меньше
Aнализ частот аллелей ряда генетических маркеров у алтайцев и ряда народов Сибири
Eщё в 1999 г. А.Ф. Назарова провела анализ частот аллелей ряда генетических маркеров у алтайцев и ряда народов Сибири, и показала, что эти частоты можно группировать как «европеоидные», «монголоидные» и промежуточные между ними. Например, генетические частоты по локусу GC2 (белка, связывающего витамин D) для очевидных европеоидов шведов (0.28), шотландцев (0.29), сербов (0.30) русских (0.32), чехов (0.32), англичан (0.29), французов (0.27), итальянцев (0.26), басков (0.31), поляков (0.34), немцев (0.29) равны в средне 0.З0±0.02 данные А.Ф. Назаровой). Напротив, соответствующие частоты у явных монголоидов - якутов (0.17), монголов (0.20), ненцев (0.21), удмуртов (0.24), вьетнамцев (0.20),чукчей (0.22), эскимосов Америки (0.17), манси (0.19), японцев (0.23), китайцев (0.24), лопарей (0.23) равны в средне 0.21 ±0.03. юда же попадают финны - 0.22. В то же время для алтайцев этот показатель раве 0.26. Что похоже на переходную форму.
Ещё пример - генетические частоты по локусу НР1 (гетерохроматинового белка 1) для тех же групп - шведов (0.40), шотландцев (0.36), сербов (0.39) русских (0.39), чехов (0.36), англичан (0.41), французов (0.39), итальянцев (0.37), басков (0.44), поляков (0.36), немцев (0.41) равны в средне 0.39±0.03 данные А.Ф. Назаровой), в то время как соответствующие частоты у монголоидов - якутов (0.32), монголов (0.29), ненцев (0.26), удмуртов (0.27), вьетнамцев (0.29),чукчей (0.31), эскимосов Америки (0.35), манси (0.41), японцев (0.24), китайцев (0.30), лопарей (0.59) равны в средне 0.30±0.05 без лопарей) или 0.33±0.10 (с лопарями). Финны по этому показателю (0.36) тяготеют скорее к «европеоидным» частотам. А у алтайцев 0.34, опять переходная форма.
Как видно из приведенных данных, «европеоидность» или «монголоидность» частот по отдельных локусам может иметь место, но попадаются исключения, свидетельствующие или о «смазанности» показателей (ведь эти показатели брались у современных людей, порой с перемешанными генами разных популяций за прошедшие тысячелетия и десятки тысяч лет), или о тривиальных погрешностях определений, которые никогда нельзя исключать полностью.
Кстати, следует отметить, что в этом контексте гаплогруппа Y-хромосомы является «бинарной» - либо она одна, либо другая и сохраняется десятки тысяч лет без изменений. Нет и не может быть смешанных гаплогрупп Y-хромосомы. Это -метка навечно и истребляется только со своим носителем. В этом смысле гаплогруппа Y-хромосомы всегда сопровождает потомков древнейших людей и передвигается вместе с ними. Она может вытесняться другой гаплогруппой, но тогда становится древнейшей меткой той, другой гаплогруппы. С другой стороны, она не отражает именно перемешиваемости генов, их сегодняшнего состояния, причём как мужчин, так и женщин, как отражают генетические частоты. Как мы видим, информация по генетическим частотам и по гаплогруппам-гаплотипам Y-хромосомы является взаимно дополняющей.
Именно потому, что отдельные локусы могут давать «смазанную» информацию, смазанную постепенным стиранием расовых и других различий, в литературе разработан метод «генетических расстояний», в котором частоты по отдельным локусам суммируются, усредняются, и приводятся к единой шкале [Nei, 1971, 1972, 1973, 1974, 1977, 1978; Nozawa et al, 1982; Ward and Skribinski, 1985; Cavalli-Sforza et al, 1971, 1988]. Это даёт показатель под названием «генетическое расстояние» между популяциями, и принимается за меру временной дистанции от их общего предка.
В работах А.Ф. Назаровой была создана матрица генетических расстояний, вычисленных по 28 аллелям двенадцати локусов белков, ферментов и групп крови [Назарова, 1999, 2002, 2005; Nazarova, 2006, 2008, 2009a, 2009b], и расчёт генетических расстояний проводился по методу [Cavalli-Sforza and Bodmer, 1971], в котором генетические расстояния рассчитываются как «угол расхождения популяций», в виде усреднённого отношения функции суммы «генетических расстояний» по каждому локусу и суммарному числу аллелей в локусах. Эти данные были рассчитаны для 11, и далее 55 популяций Европы, Азии и Америки.
Oтметим, что на основании полученных данных А.Ф. Назарова ещё в 1999 году выдвинула гипотезу о происхождении европеоидов, северных монголоидов и американских индейцев из одной древней пра-популяции, озаглавив свою статью «Гeнетические данные к проблеме дифференциации северных монголоидов, европеоидов и америндов на территории Евразии» [Назарова, 1999]. Эти же данные и выводы суммированы в её книге «Генетический портрет народов мира» (М., Липецкое изд-во, 2010), в главе «Популяции, переходные между монголоидами и европеоидами, и возможный путь формирования европеоидов».
В подтверждение своей концепции А.Ф. Назарова привлекает литературные данные о полиморфизме митохонд-риальной ДНК у народов Евразии и американских индейцев, а также данные палеогеографии, палеолингвистики и палеозоологии. Как видно из данных приведённых выше в настоящем разделе, эта гипотеза через 10 лет блестяще подтвердилась с помощью совершенно независимых данных, основанных на анализе гаплогрупп Y-хромосомы, а именно сводной гаплогруппы NOP, из которой произошли и монголоиды (гаплогруппа N), и европеоиды (гаплогруппа R), и американские индейцы (гаплогруппа Q).
Видимо, типичными представителями самых древних европеоидов являются скандинавы - шведы, норвежцы, датчане, исландцы, у которых к тому же преобладает гаплогруппа I1 (48%, 42%, 34% и 33%, соответственно). Это - северные европеоиды. Свидетельством размывания расовых признаков является немалая доля этой гаплогруппы у финнов - от древних 100% гаплогруппы N у них осталось (по регионам) от 62 до 77% N, и уже четверть Y-хромосомного пула заняла гаплогруппа I1 (20 - 26%). Иначе говоря, у финнов идёт заметный дрейф от монголоидной гаплогруппы N.
Список родственников русских:
Лужичане, Поляки, Украинцы, Белорусы, Венгры, Чехи, Словаки, Латыши, Литовцы, Киргизы, Алтайцы, Таджики, Ишкашим, Белуджи, Пенджабцы, Пуштуны, Талыши, Норвежцы,Немцы, Австрийцы, Швейцарцы, Поляки, Боснийцы, Литовцы, Ирландцы, Шотландцы, Иранцы, Коми, Чуваши, Удмурты, Ненцы,Осетины, Азербайджанцы.То что русские родственники киргизов, таджиков, можно не сомневаться - вот например Дерево родства 61 евразийской популяции основанное Y-хромосоме. :))
Смотри
http://www.pnas.org/content/98/18/10244.full
Одонтология
Морфогенез одонтологических признаков обусловлен главным образом генетическими факторами и слабо зависит от влияния внешней среды. Полиморфизм зубочелюстной системы широко используется в исследованиях проблем этногенеза.
Некоторые одонтологические характеристики в различных группах русских (%):
| Группа | Лопатообразность резцов (баллы 2-3) | Сумма четырёхбугорковых М1 | Сумма шестибугорковых М1 | Сумма пятибугорковых М2 | 2med (II) |
|---|---|---|---|---|---|
| Кардымово (Смоленская область) | 7.4 | 6.3 | 3.5 | 14.6 | 40.0 |
| Починок (Смоленская область) | 9.4 | 18.6 | 3.0 | 13.7 | 50.5 |
| Смоленск | 8.9 | 12.9 | 5.5 | 10.8 | 42.6 |
| Старый Изборск (Псковская область) | 10.1 | 16.4 | 1.5 | 13.1 | 45.6 |
| Псков | 8.0 | 11.4 | 2.0 | 5.4 | 50.6 |
| Луга (Ленинградская область) | 2.4 | 8.8 | 3.3 | 12.6 | 65.4 |
| Мста (Новгородская область) | 4.7 | 18.6 | 1.4 | 5.6 | 54.1 |
| Валдай (Новгородская область) | 4.8 | 15.0 | 1.3 | 9.3 | 44.3 |
| Новгород | 5.6 | 16.9 | 1.1 | 8.2 | 58.1 |
| Западная Двина (Калининская область) | 8.5 | 6.5 | 2.2 | 8.1 | 57.8 |
| Калязин (Калининская область) | 4.4 | 11.6 | 3.5 | 6.6 | 46.3 |
| Вологодская область | 2.2 | 9.0 | 1.0 | 13.0 | 46.5 |
| Архангельская область | 9.3 | 20.5 | 2.0 | 15.3 | 34.1 |
| Рязанская область | 3.6 | 6.9 | 3.0 | 11.6 | 51.6 |
| Тамбовская область | 4.6 | 7.0 | 2.0 | 14.3 | 43.5 |
| Липецкая область | 5.2 | 13.6 | 2.3 | 8.3 | 45.3 |
| Воронежская область | 5.3 | 9.0 | 1.1 | 7.5 | 41.2 |
| Ростовская область | 7.4 | 6.9 | 4.6 | 18.3 | 40.0 |
| Тульская область | 3.3 | 10.0 | 2.2 | 10.5 | 56.5 |
| Орловская область | 1.7 | 8.4 | 1.1 | 17.6 | 47.4 |
| Курская область | 2.3 | 5.0 | 6.2 | 18.1 | 27.6 |
| Белгородская область | 1.7 | 3.0 | 5.0 | 7.1 | 42.4 |
| Калужская область | 8.3 | 10.1 | 3.8 | 6.6 | 37.7 |
| Брянская область | 10.7 | 16.9 | 2.4 | 16.4 | 30.0 |
Значения основных признаков одонтологии однозначно характеризуют русских как представителей западного круга форм. Одонтологические показатели русского населения наиболее близки к показателям среднеевропейского одонтологического типа.
Межгрупповые различия признаков у русских значительно меньше, чем между русскими и другими народами, что позволяет говорить о единстве русских. Тем не менее, на основании этих различий были выделенны 2 основных одонтологических комплекса: северо-западный и южный, связанные между собой рядом переходных форм. Одонтологические типы в составе русского народа напрямую связаны с типами Восточной Европы в целом: среднеевропейским и северным грацильным. Выявлено, что северо-западный тип русских сформировался под влиянием северного грацильного типа Восточной Европы. В частности, для северо-западного одонтологического типа русских характерно некоторое повышение встречаемости лопатообразных резцов, что сближает эти популяции с соседними восточными латышами, эстонцами, финнами, карелами, вепсами и может объясняться влиянием общего субстрата. В целом встречаемость лопатообразных резцов, характерных для монголоидных популяций, у русских повсеместно низка.
Достоверных различий между городским и сельским населением областей, где такие исследования проводились, не выявлено.
Дерматоглифика

Показано, что дерматоглифические признаки не подвержены влиянию направленного отбора и могут быть использованы в популяционных исследованиях наравне с генетическими.
Русские популяции являются достаточно однородными по признакам кожного рельефа. Тем не менее, в северных популяциях отмечаются черты северных европеоидов (пониженный дельтовый индекс сочетается с низким процентом добавочных межпальцевых трирадиусов), на юге — южноевропеоидные (относительно высокий процент добавочных межпальцевых трирадиусов сочетается с пониженной частотой осевого проксимального трирадиуса ладони у мужчин и относительно высоким дельтовым индексом у женщин).
Европеоидно-монголоидный комплекс у русских, как правило, невысокий и в среднем составляет 41,9 у мужчин и 44,2 у женщин.
Восточные славяне обнаруживают высокое сходство друг с другом по дерматоглифическим данным. Ключевые признаки кожного рельефа у восточных славян варьируют незначительно, коэффициенты их вариации меньше, чем у европеоидов Европы и на территории Восточной Европы в целом. Дерматоглифический тип восточных славян является характерным для европеоидов Восточной Европы.
Минимальные обобщённые дерматоглифические расстояния отделяют восточных славян от мордвы, немцев, коми, молдаван, шведов. Обобщённые дерматоглифические расстояния между восточными славянами и литовцами, латышами,эстонцами, финнами, казанскими татарами, гагаузами выше, однако также находятся в пределах малых расстояний.
Генетика

Генофонд русского народа — типично европейский. Процент восточноевразийских генетических маркеров в составе русского генофонда не превышает средние показатели по Европе. Результаты изучения русского генофонда показали его близость с населением практически всей Европы, при этом выявлены значительные отличия от населения Урала и Кавказа.
Степень гетерогенности русского генофонда выше, чем у большинства остальных народов Европы. Это может быть связано с большой численностью русских и большой территорией расселения. Следует отметить, что население Восточной Европы в целом более гетерогенно, чем население Западной Европы. При этом населениеЗападной Сибири, наоборот, более гетерогенно, чем русские и иные народы Восточной Европы. Таким образом генофонды Восточной Европы, в том числе и русский генофонд по уровню гетерогенности занимают промежуточное положение между Западной Европой и генофондами народов Западной Сибири. В пределах самого русского генофонда можно отметить значительные отличия между северными популяциями русских, и значительно меньшие между южными и центральными, то есть население Русского Севера более гетерогенно.
Для русского генофонда выявлена широтная изменчивость (изменение характеристик по оси север — юг). Этим русский генофонд отличается от общего генофондаВосточной Европы, для которого характерна долготная изменчивость. Таким образом, характер изменчивости русского генофонда не сводим к исходной изменчивостисубстратов, на основе которых он формировался. Предполагается, что широтная изменчивость русского генофонда возникла или усилилась в ходе собственной историирусского народа.
Маркеры Y-хромосомы ДНК
Распределение гаплогрупп Y-хромосомы в различных русских популяциях:
| Популяция | N= | C | E1b1b1 | E3b3 | F* | G1* | G2 | I* | I1 | I2 | I1c | J1 | J2 | K* | K2 | N* | N1b | N1c | Q | R1a | R1b2 | R1b3 | R2 |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| M130 | M78 | M123 | M89 | M285 | P15 | M170 | M253 | P37 | M223 | M267 | M172 | M9 | M70 | M231 | P43 | TAT | M242 | SRY1532 | M73 | M269 | M124 | ||
| Мезень | 54 | 0 | 0 | 0 | 1.9 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 7.4 | 46.3 | 0 | 44.4 | 0 | 0 | 0 |
| Пинега | 114 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0.9 | 4.4 | 0 | 0.9 | 0 | 0 | 0 | 0 | 15.8 | 23.7 | 0.9 | 39.5 | 0 | 14 | 0 |
| Красноборск | 91 | 0 | 0 | 0 | 1.1 | 0 | 2.2 | 3.3 | 12.1 | 9.9 | 0 | 0 | 5.5 | 0 | 0 | 0 | 3.3 | 36.3 | 0 | 19.8 | 0 | 6.6 | 0 |
| Вологда | 121 | 0 | 0.8 | 0 | 0 | 1.7 | 0.8 | 0 | 11.6 | 8.3 | 1.7 | 0 | 0.8 | 0 | 0 | 0 | 3.3 | 35.5 | 1.7 | 33.1 | 0 | 0.8 | 0 |
| Унжа | 52 | 0 | 5.8 | 0 | 1.9 | 0 | 0 | 0 | 11.5 | 11.5 | 3.8 | 3.8 | 3.8 | 0 | 0 | 0 | 0 | 13.5 | 0 | 32.7 | 1.9 | 9.6 | 0 |
| Кашин | 73 | 0 | 4.1 | 0 | 1.4 | 0 | 0 | 0 | 2.7 | 8.2 | 4.1 | 0 | 4.1 | 0 | 0 | 0 | 0 | 11 | 1.4 | 56.2 | 0 | 6.8 | 0 |
| Порхов | 57 | 0 | 1.8 | 1.8 | 0 | 0 | 0 | 1.8 | 3.5 | 10.5 | 0 | 0 | 0 | 3.5 | 0 | 1.8 | 1.8 | 15.8 | 0 | 52.6 | 0 | 5.3 | 0 |
| Остров | 75 | 0 | 4 | 0 | 0 | 0 | 0 | 1.3 | 6.7 | 9.3 | 0 | 1.3 | 1.3 | 0 | 0 | 0 | 0 | 28 | 0 | 45.3 | 0 | 2.7 | 0 |
| Рославль | 107 | 0.9 | 7.5 | 0 | 0.9 | 0 | 0 | 0 | 1.9 | 10.3 | 0 | 0 | 2.8 | 1.9 | 2.8 | 0 | 0.9 | 13.1 | 0 | 45.8 | 0 | 11.2 | 0 |
| Ливны | 110 | 0 | 0.9 | 0 | 0 | 0 | 0 | 0 | 8.2 | 13.6 | 0.9 | 0 | 0.9 | 0 | 2.7 | 0 | 0.9 | 4.5 | 0.9 | 62.7 | 0 | 3.6 | 0 |
| Пристень | 45 | 0 | 2.2 | 0 | 0 | 0 | 0 | 0 | 4.4 | 17.8 | 0 | 0 | 2.2 | 0 | 2.2 | 0 | 0 | 13.3 | 0 | 55.6 | 0 | 2.2 | 0 |
| Репьёвка | 96 | 0 | 1 | 0 | 1 | 0 | 2.1 | 1 | 3.1 | 16.7 | 0 | 2.1 | 1 | 0 | 0 | 0 | 0 | 6.3 | 0 | 59.4 | 0 | 5.2 | 1 |
| Белгород | 143 | 0.7 | 0.7 | 0 | 0 | 0 | 0.7 | 0 | 3.5 | 12.6 | 0.7 | 0 | 4.2 | 0.7 | 1.4 | 0 | 0.7 | 11.9 | 0 | 59.4 | 0 | 2.8 | 0 |
| Кубанские казаки | 90 | 0 | 3.3 | 0 | 0 | 0 | 1.1 | 1.1 | 4.4 | 16.5 | 2.2 | 0 | 4.4 | 0 | 1.1 | 0 | 1.1 | 6.6 | 1.1 | 47.3 | 0 | 8.8 | 0 |
По результатам исследования Y-хромосомы русские на 99,7 % имеют происхождение из Западной Евразии.
Генетический пул русских сходен с остальными популяциями центральной и восточной Европы. В пределах русского генетического пула чётко выделяются две группы популяций. В северной группе (Мезень, Пинега, Красноборск) обнаруживается близость с соседними балтскими и финно-угорскими популяциями. Южно-центральная группа, к которой относится подавляющее большинство русских популяций, входит в общий кластер с белорусами, украинцами и поляками.
Маркеры митохондриальной ДНК
Распределение гаплогрупп митохондриальной ДНК в различных русских популяциях:
| Популяция | N= | H | V | Pre*V | HV* | R1 | J | T* | T1 | K | U1 | U2 | U3 | U4 | U5a | U6b | U7 | U8 | I | N1a | N1b | W | X | C | D | G2a | M1 | L1b | L3b |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Тула | 73 | 58.9 | 1.4 | 1.4 | 1.4 | 0 | 5.5 | 8.2 | 1.4 | 2.7 | 0 | 0 | 2.7 | 4.1 | 2.7 | 2.7 | 0 | 0 | 2.7 | 1.4 | 1.4 | 0 | 0 | 0 | 0 | 0 | 0 | 1.4 | 0 |
| Калуга | 71 | 38.0 | 2.8 | 0 | 2.8 | 0 | 4.2 | 11.3 | 2.8 | 4.2 | 1.4 | 2.8 | 1.4 | 8.5 | 8.5 | 2.8 | 0 | 0 | 1.4 | 0 | 0 | 0 | 2.8 | 2.8 | 0 | 0 | 0 | 0 | 1.4 |
| Владимир | 72 | 38.5 | 0 | 0 | 1.4 | 0 | 8.3 | 8.3 | 11.1 | 6.9 | 0 | 1.4 | 1.4 | 2.8 | 6.9 | 0 | 1.4 | 0 | 5.6 | 1.4 | 1.4 | 4.2 | 1.4 | 0 | 0 | 0 | 1.4 | 0 | 0 |
| Ярославль | 41 | 48.8 | 0 | 2.4 | 4.9 | 0 | 4.9 | 2.4 | 0 | 0 | 7.3 | 0 | 0 | 0 | 4.9 | 9.8 | 0 | 0 | 0 | 2.4 | 2.4 | 2.4 | 0 | 4.9 | 0 | 2.4 | 0 | 0 | 0 |
| Псков | 68 | 42.6 | 5.9 | 0 | 0 | 1.5 | 11.8 | 2.9 | 2.9 | 0 | 0 | 2.9 | 0 | 2.9 | 17.6 | 2.9 | 0 | 1.5 | 0 | 0 | 0 | 2.9 | 0 | 0 | 0 | 1.5 | 0 | 0 | 0 |
По результатам исследования мтДНК русские в среднем на 97,9 % имеют происхождение из Западной Евразии. В одной из работ (2002) приводилась также цифра 98,5 %. Существуют некоторые различия в доле восточноевразийского компонента в генофонде различных русских популяций. Среди русских южных и центральных регионов доля восточноевразийских гаплогрупп митохондриальной ДНК составляет 1,3 %, что сравнимо с показателями западных славян (1,6 %), среди северных русских до 4 %, среди поморов до 5 %.
По результатам исследования мтДНК, можно выделить две группы русских популяций: юго-западную и северо-восточную. Обе группы обнаруживают большое сходство с европейскими популяциями.
Аутосомные маркеры
Аутосомные маркеры — маркеры неполовых хромосом. Как правило, эти гены кодируют синтез определённых белков, ферментов и т. п.
Биохимические маркеры
Генетико-биохимические маркеры, как правило, представляют собой гены, кодирующие синтез ферментов. Полиморфизм некоторых иммунологических маркеров крови также может быть отнесён к биохимическому полиморфизму, так как эти гены кодируют ферменты, катализирующие соединение соответствующих сахаров с группоспецифическими гликопротеидами или гликолипидами. Показано, что полиморфизм многих биохимических маркеров может быть использован в популяционных исследованиях.
В таблице приведены частоты некоторых биохимических маркеров эритроцитарной фракции крови, сыворотки крови, иммунологических маркеров крови.
| Биохимический маркер | Геногеография | Аллельные частоты у русских | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Группа крови (система AB0) | Частота гена А — 0,05 среди населения Центральной и Южной Америки, 0,25—0,3 среди населения Центральной Европы, 0,4—0,45 среди населения Центральной Австралии и Северной Европы. Частота гена В — 0,0—0,05 среди населения американского континента, 0,05—0,1 среди населения Западной и Центральной Европы, 0,1—0,2 среди населения Восточной Европы, 0,2—0,3 среди азиатского населения. Частота гена 0 — 0,5—0,55 среди населения некоторых азиатских регионов, 0,6—0,65 среди населения Центральной и Восточной Европы, 0,65—0,7 среди населения Южной и Западной Европы, 0,95—1,0 среди населения Центральной и Южной Америки. | 0 | A | B | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 0.598 | 0.247 | 0.156 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Резус-фактор | В странах Тихоокеанского бассейна (Америка, Австралия, Океания, Восточная и Юго-Восточная Азия) концентрация варианта D, как правило, выше 0.8, в Южной и Юго-Западной Азии, а также в Африке(южнее Сахары) до 0.8, в Европе и Северной Африке она ещё ниже, падая у берегов Атлантики (Испания, Франция, Англия, Скандинавия) ниже 0.6. | Rh*D | Rh*d | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 0.632 | 0.368 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Кислая эритроцитарная фосфатаза | Обнаружено шесть основных фенотипов, определяемых аллелями АСР1*А, АСР1*В, АСР1*С. Из них аллель АСР1*С является самым редким и встречается у европеоидов с частотой 0,03—0,07, у монголоидов, негроидов и аборигенов Австралии с частотой ещё меньшей, а в некоторых популяциях отсутствует полностью. | ACP1*A | ACP1*B | ACP1*C | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 0.339 | 0.608 | 0.053 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Глиоксалаза 1 | Два аллеля GLO1*1 и GLO1*2 по крайней мере среди европейского населения распределяются почти с одинаковой частотой. | GLO1*1 | GLO1*2 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 0.327 | 0.673 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Трансферрин | Два наиболее распространённых аллеля TF*C1 и TF*C2 присутствуют во всех популяциях. У европейцев и американских белых TF*C1 встречается с частотой 75—78 %. TF*C2 в европейских популяциях имеет частоту 13—19 %, в азиатских 15—34 %. TF*C3 встречается европеоидных группах с частотой 4—7 %, в популяциях Индии — 1—4 %, отдельные случаи встречаемости в Восточной Азии, некоторых группах Тихоокеанского бассейна и у американских негров. В других популяциях он полностью отсутствует. По мнению ряда авторов, аллель С3 имеет европейское происхождение. TF*DChi является маркером монголоидных популяций и он широко распространён в Восточной Азии. | TF*B | TF*C1 | TF*C2 | TF*C3 | TF*DChi | |||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 0.018 | 0.827 | 0.112 | 0.037 | 0.005 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Группоспецифический компонент | В европейских популяциях соотношение частот встречаемости аллелей GC*1F:GC*1S составляет от 2:10 до 3:10. Среди монголоидных популяций в отличие от европейских частота встречаемости GC*1F преобладает, ещё большая плотность этого гена свойственна негроидным популяциям. Частота встречаемости аллеля GC*2 составляет среди европеоидных популяций около 28 %,монголоидных — около 23 %, негроидных — около 8 % и среди аборигенов Австралии — около 12 %. | GC*1F | GC*1S | GC*2 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 0.119 | 0.615 | 0.262 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Фосфоглюкомутаза | Обнаружена направленная географическая изменчивость частот фосфоглюкомутазы в зависимости от климатических характеристик. | PGM1*1+ | PGM1*1- | PGM1*2+ | PGM1*2- | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 0.591 | 0.241 | 0.090 | 0.079 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Эстераза D | Самыми высокими значениями аллеля EstD*1 характеризуются негроидные группы (0.89—0.95). Несколько ниже концентрация гена EstD*1 у европейцев (0.85—0.89). Таким образом, по уменьшению частоты гена EstD*1 популяции мира располагаются в следующем порядке: негроиды, европеоиды, группы Индии, монголоиды. Наиболее высокие частоты гена EstD*2 присущи монголоидам (до 0.408 у тибетцев). Остальные аллели встречаются чрезвычайно редко. | EstD*1 | EstD*2 | EstD*5 | EstD*7 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 0.885 | 0.112 | 0.002 | 0.001 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Минисателлитные маркеры

Наиболее изученными в российских популяциях гипервариабельными минисателлитными локусами являются D1S80 , D1S111, IL-1RN и 3’ApoB, так как они представляют наибольший интерес с точки зрения судебно-генетической экспертизы. Изучение полиморфизма локусов D1S80 и 3’ApoB также используется в популяционных исследованиях, так как данные маркеры обладают весьма высокой дифференцирующей способностью. Минисателлитный локус D1S80 локализован на коротком плече 1-й хромосомы. Минисателлитный локус 3’АроВ локализован на коротком плече 2-й хромосомы и представляет собой высокополиморфный участок на расстоянии 500 пар оснований после 3'-конца гена аполипопротеина В.
По результатам исследований минисателлитных локусов D1S80 и 3’ApoB можно отметить высокое единство всех восточнославянских популяций и их значительные отличия от соседних финно-угорских, тюркских и северокавказских народов, что может объясняться общим происхождением восточных славян и незначительностью влияния на их генофонд соседних этносов.
Главный комплекс гистосовместимости
Комплекс генов HLA (главного комплекса гистосовместимости) локализован на коротком плече 6-й хромосомы и занимает 3500 тысяч пар оснований, содержит более 220 генов. Он выполняет функции генетического контроля иммунного ответа и поддержания иммунного гомеостаза. Гены локуса HLA DP, DQ и DR кодируют молекулы II класса, выявленные на поверхности антиген-представляющих клеток (хелперов), в частности, В-лимфоцитов, Т-хелперов и некоторых других.
Система HLA является одной из наиболее полиморфных генетических систем. Показано, что, несмотря на влияние направленного естественного отбора, распределение генов HLA II класса адекватно отражает геногеографию этнических групп, и особенности полиморфизма генов HLA могут быть использованы в популяционных исследованиях.
Наибольший полиморфизм проявляет ген DRB1, его изучение наиболее информативно при популяционных исследованиях.
Частоты аллелей системы DRB1 в некоторых восточнославянских популяциях:
| Популяция | Северные белорусы (Витебская область) | Южные белорусы (Брестская область) | Восточные украинцы (Хмельницкая область) | Западные украинцы (Львовская область) | Русские (Вологодская область) |
|---|---|---|---|---|---|
| Аллель DRB1 | N=100 | N=105 | N=137 | N=102 | N=121 |
| *01 | 10.0 % | 10.5 % | 9.5 % | 12.2 % | 12.4 % |
| *03 | 7.0 % | 9.0 % | 5.5 % | 7.8 % | 7.4 % |
| *04 | 7.5 % | 12.9 % | 10.9 % | 11.3 % | 14.0 |
| *07 | 10.5 | 10.5 | 14.6 | 15.2 | 14.9 |
| *08 | 5.5 % | 2.9 % | 3.6 % | 5.4 % | 5.4 % |
| *09 | 0 | 1.9 % | 0.4 % | 1.5 % | 2.5 % |
| *10 | 0 | 0.5 % | 1.1 % | 0 | 0.4 % |
| *11 | 15.0 % | 13.3 % | 18.6 % | 18.6 % | 9.5 % |
| *12 | 4.0 % | 3.8 % | 2.2 % | 0.5 % | 1.2 % |
| *13 | 16.5 % | 11.9 % | 12.4 % | 10.8 % | 14.9 % |
| *14 | 1.0 % | 2.9 % | 2.2 % | 2.0 % | 1.2 % |
| *15 | 18.0 % | 13.3 % | 10.2 % | 9.8 % | 14.5 % |
| *16 | 5.0 % | 6.7 % | 8.8 % | 5.9 % | 1.6 % |
Частоты аллелей гена DRB1 в популяциях восточных славян соответствует типичному западноевразийскому варианту распределения. Встречаемость частот аллелей DRB1*09 и DRB1*12, характерных для монголоидных популяций, у русских низка.
По результатам многомерного анализа полиморфизма системы HLA в популяциях Европы выделяется три кластера: североевропейский, центральноевропейский и южноевропейский. Русские входят в североевропейский кластер, наряду с немцами, датчанами, голландцами, финнами, ирландцами и англичанами.
Смотри;http://ru.wikipedia.org/wiki/%C0%ED%F2%F0%EE%EF%EE%EB%EE%E3%E8%FF_%F0%F3%F1%F1%EA%E8%F5
У всех голубоглазых людей был один общий предок
Различные фенотипы цвета глаз (a-d). Людей с голубыми глазами без единого пятнышка (a) относят к генотипу rs12913832 G/G, в то время как всех остальных (b-d) к генотипу rs12913832 A/G (фото Københavns Universitet).
Команда учёных под руководством доцента Ханса Эиберга (Hans Eiberg) из университета Копенгагена (Københavns Universitet) изучила мутацию, которая произошла около 6-10 тысяч лет назад, и обнаружила, что у ныне живущих людей с голубыми глазами, вероятно, был общий предок.
Произошедшая около 10 тысяч лет назад мутация, которая затронула ген OCA2, привела к тому, что производство меланина в радужной оболочке глаза было ограничено (и коричневая окраска была «разбавлена» до голубой).
Если бы OCA2 был бы полностью выключен, человек стал бы альбиносом, так как прекратилась бы выработка меланина в волосах, глазах и коже. В данном случае изменения привели лишь к частичному ограничению его производства организмом.
И хотя цвет глаз определяется различными параметрами, а не только количеством меланина, у голубоглазых его содержание в радужке значительно меньше, чем у всех остальных людей.
Изучая ДНК митохондрий, учёные установили, что у всех людей с голубым цветом глаз присутствует незначительное изменение в определённом месте двойной спирали. В то время как у кареглазых нет какой-то одной характерной «точки», их вариации в строении ДНК (в той её области, что отвечает за процессы, происходящие с меланином в организме) более обширны.
Из этого учёные и сделали вывод о том, что у всех голубоглазых был один общий предок, у которого и произошла данная мутация.
http://www.membrana.ru/particle/12269
Структурный анализ северо-востока Европы
Любопытные новости от проекта Eurogenes. Ниже приведены результаты структурного анализа 400 образцов ДНК (65000 SNP) жителей стран Северо-Восточной Европы, для контекста дополненных средиземноморскими, кавказскими и сибирскими популяциями. Все четверо дедушек и бабушек носителей ДНК происходили из одной страны. Массив исходных данных был распределён по 6 кластерам, соотношение которых в отдельных популяциях и представляет интерес.
Региональное расположение кластеров:
— жёлтый — Западная Азия;
— розовый — Средиземноморье;
— зелёный — Северная Европа;
— синий — Южная Балтика;
— голубой — Волга-Урал;
— красный — Сибирь.
Таблица расстояний между кластерами:
Доля зелёного «северо-европейского» компонента достигает максимума в Скандинавии и Финляндии, в то время как среди остальных популяций он ниже в полтора раза, а у чувашей отсутствует полностью. Синий «южно-балтийский» компонент распределён схожим образом, но менее широко: он встречается у славян, балтов и венгров, но практически отсутствует в Скандинавии, среди финнов и чувашей. Голубой компонент «Волга-Урал» доминирует у чувашей, высок у русских из Вологды, прибалтийских и поволжских финнов, ниже у остальных русских и тонко распределён по другим северо-восточным европейцам. Расстояния от волго-уральского кластера до двух других меньше (0,0017 и 0,0020), чем между южно-балтийским и северо-европейским кластерами (0,0025).
Все три кластера ассоциируются с Северо-Восточной Европой, явно имеют общее происхождение и отражают процесс формирования облика современного северо-европейского населения. Тёмно синий «южно-балтийский» компонент здесь отчётливо связан с расселением славян и балтов по северо-востоку континентальной Европы (культуры шнуровой керамики?), нёсших поровну зелёного и характерного синего. Голубой «волго-уральский» компонент — это, вероятно, архаичный европеоидный субстрат, сохранивший свои позиции в северо-восточных областях континента как в качестве предков как не связанного с финнами населения (поздние восточные потомкисвидерской культуры: нарвская, верхневолжская, неманская?), так и предков прибалтийских финнов (финноязычные народы формировались на европеоидной основе). В континентальной части это население испытало влияние балтов/славян, местами с последующим возвратом к финской культуре: структура поволжских финнов наполовину южно-балтийская, наполовину — финская.
Что касается зелёного «северо-европейского» кластера, можно согласиться с Дэвидом, что в него вошли все те автохтонные в Северной Европе аллели, которые не попали в «южно-балтийский» и «волго-уральский» кластеры. Он не демонстрирует корреляции с германскими языками и не выделяется как изолированный от двух своих северных братьев компонент, но в целом показывает бóльшую близость к более архаичному «волго-уральскому» кластеру. Вероятно, он охватывает всё то северо-европеоидное население, которое широко расселилось по северу Европы к моменту начала экспансии предков балтов и славян, но не было затронуто влиянием последних (в Скандинавии — потомки аренсбургской культуры и последующих?).
Если рассматривать эти данные генетики с точки зрения антропологии и археологии, то они не открывают ничего нового, укладываются в непротиворечивую, давно известную картину. В действительности, они рассказывают историю Европы с собственной точки зрения и акцентируют внимание на том, что важно для популяционной генетики как таковой. В частности, мы видим отчётливое указание, что к началу формирования на континенте балтов и славян в Скандинавии и Финляндии уже жило родственное им северо-европеоидное население, а у них обоих были ещё одни, более древние предки.
http://drfaust-spb.livejournal.com/261396.html
Аутосомные ДНК о происхождении славян и балтов
Вернёмся к последней диаграмме структурного анализа ДНК северо-востока Европы, чтобы извлечь из неё ещё не извлечённые следствия.
Немного общих мест. Популяционная генетика примечательна тем, что рассказывает историю народов на языке непосредственно данных о населении, без обращения к производным от населения: археологии и лингвистике. Особенность аутосомных ДНК в том, что они показывают, чьим именно потомком является тот или иной современный человек, — за счёт равного наследования структуры ДНК от обоих родителей из поколения в поколение (в отличие от данных однородительских маркёров, которые позволяют выделить дальних предков человека по материнской и отцовской линиям). Другими словами, результат структурного анализа аутосомных ДНК указывает, откуда пришли предки человека, пропорционально их удельному весу.
Взглянем на распределение «южнобалтийского» компонента в европейских популяциях. Этот компонент выделяется как половина синего и половина зелёного. Он объединяет балтов со славянами и доминирует среди литовцев.К западу от Литвы увеличивается южноевропейское влияние, к востоку видно влияние популяций Сибири и — слабее — той же Южной Европы. Среди народов Скандинавии, финнов и чувашей южнобалтийский компонент или невелик, или отсутствует. При этом и среди западных славян, и среди восточных встречаются отдельные образцы, структурно идентичные литовцам.
Всё перечисленное указывает на то, что южнобалтийский компонент формировался в континентальной зоне Северо-Востока Европы, включавшей территории от Польши до России, от Балтики до Венгрии, и на исходном этапе не имел южноевропейских или сибирских элементов. В последующее время популяции с территории Польши, Украины и Белоруссии получают южноевропейское влияние.
Из диаграммы невозможно однозначно заключить, какова была история появления южнобалтийского компонента на территории России: был ли он здесь автохтонным с последующим приходом населения, обладавшим структурой современных прибалтийских финнов, или пришёл поверх последнего. Известно только, что в структуре финнов (и прибалтийских, и поволжских) доминируют более древние северо-европеоидные компоненты (зелёный, голубой). В любом случае, на территории России южнобалтийский компонент появляется дважды: в первый раз — сам по себе (идентичные или близкие к литовцам русские образцы), во второй раз — совместно с незначительным южноевропейским влиянием (по образцу белорусов и украинцев).
Резюмируя. Об истории «южнобалтийского» компонента известно, что он:
1. близок к компонентам более архаичных северных европейцев (зелёный, голубой);
2. сформировался в лесной зоне до прихода туда компонентов Южной Европы (то есть до середины 6 тыс. до н.э.);
3. фиксируется на территории России дважды: сначала — сам по себе, позже — совместно с незначительным южноевропейским влиянием (жёлтый, розовый).
Вопрос параллелей данных генетики с историческими балтами и славянами можно решить простейшим образом: приписать перенос чистого южнобалтийского компонента балтам, перенос его же с незначительным южноевропеоидным влиянием — славянам, и забыть о мезолитическом субстрате, списав его на финнов. Но такой произвол — плохое решение. Фактически мы видим высокую однородность континентального населения северо-востока Европы и всего одну волну миграции на восток из Центральной Европы, которая была метисирована с южными европейцами, — та волна почти без следа растворилась в местном населении (сравнить уровни южного влияния у поляков, украинцев, белорусов и русских; у русских из Вологды жёлтый цвет отсутствует в принципе).
Это означает, что роль миграций в формировании славян и балтов сильно переоценена: процессы образования языков, сложения материальной культуры и переноса технологий сопровождались переносом не настолько значительных масс населения, как привыкли считать археологи и лингвисты. Картина выглядит так, как если бы развивался большой автохтонный массив населения, в котором кристаллизовались, расширялись и поглощались отдельные центры, а весь процесс катализировался влиянием населения Центральной Европы — и это влияние, безусловно, имело место.
В таком контексте разговоры о «приходе славян» неуместны, а гипотезы о происхождении славян из Южной Европы — абсолютно нелепы.
http://drfaust-spb.livejournal.com/274301.html
Всё перечисленное указывает на то, что южнобалтийский компонент формировался в континентальной зоне Северо-Востока Европы, включавшей территории от Польши до России, от Балтики до Венгрии, и на исходном этапе не имел южноевропейских или сибирских элементов. В последующее время популяции с территории Польши, Украины и Белоруссии получают южноевропейское влияние.
Из диаграммы невозможно однозначно заключить, какова была история появления южнобалтийского компонента на территории России: был ли он здесь автохтонным с последующим приходом населения, обладавшим структурой современных прибалтийских финнов, или пришёл поверх последнего. Известно только, что в структуре финнов (и прибалтийских, и поволжских) доминируют более древние северо-европеоидные компоненты (зелёный, голубой). В любом случае, на территории России южнобалтийский компонент появляется дважды: в первый раз — сам по себе (идентичные или близкие к литовцам русские образцы), во второй раз — совместно с незначительным южноевропейским влиянием (по образцу белорусов и украинцев).
Резюмируя. Об истории «южнобалтийского» компонента известно, что он:
1. близок к компонентам более архаичных северных европейцев (зелёный, голубой);
2. сформировался в лесной зоне до прихода туда компонентов Южной Европы (то есть до середины 6 тыс. до н.э.);
3. фиксируется на территории России дважды: сначала — сам по себе, позже — совместно с незначительным южноевропейским влиянием (жёлтый, розовый).
Вопрос параллелей данных генетики с историческими балтами и славянами можно решить простейшим образом: приписать перенос чистого южнобалтийского компонента балтам, перенос его же с незначительным южноевропеоидным влиянием — славянам, и забыть о мезолитическом субстрате, списав его на финнов. Но такой произвол — плохое решение. Фактически мы видим высокую однородность континентального населения северо-востока Европы и всего одну волну миграции на восток из Центральной Европы, которая была метисирована с южными европейцами, — та волна почти без следа растворилась в местном населении (сравнить уровни южного влияния у поляков, украинцев, белорусов и русских; у русских из Вологды жёлтый цвет отсутствует в принципе).
Это означает, что роль миграций в формировании славян и балтов сильно переоценена: процессы образования языков, сложения материальной культуры и переноса технологий сопровождались переносом не настолько значительных масс населения, как привыкли считать археологи и лингвисты. Картина выглядит так, как если бы развивался большой автохтонный массив населения, в котором кристаллизовались, расширялись и поглощались отдельные центры, а весь процесс катализировался влиянием населения Центральной Европы — и это влияние, безусловно, имело место.
В таком контексте разговоры о «приходе славян» неуместны, а гипотезы о происхождении славян из Южной Европы — абсолютно нелепы.
http://drfaust-spb.livejournal.com/274301.html
Анализ fastIBD Центральной и Восточной Европы.
В проекте Dodecad Ancestry Project был проведен fastIBD анализ по популяциям Центральной и Восточной Европы. Синими линиями я выделил Восточно-Европейский кластер, куда попали восточные славяне: русские, белорусы, украинцы. Зеленый квадрат выделенный красными линиями это и есть русские.
Результат анализа закономерен, русские находятся среди восточно-славянских народов: Русские, Белоруссы, Украинцы.
№ 4, 2007 г.
Борис Аркадьевич Малярчук, д. б. н., зав. лаб. генетики Ин-та биологических проблем Севера Дальневосточного отделения РАН (Магадан).© Малярчук Б.А., Деренко М.В
Мирослава Васильевна Деренко, к. б. н., вед. научн. сотр. того же института.
Немного славянской истории
Прежде чем рассказать о результатах исследования структуры русского генофонда, напомним - буквально в нескольких словах - историю славян. Эту самую многочисленную группу народов в Европе принято делить на западных славян, южных и восточных. К западным относят поляков, чехов, словаков и лужичан; к южным - болгар, сербов, хорватов, словенцев, македонцев, боснийцев, черногорцев, а к восточной - русских, украинцев и белорусов. Вполне естественно, что вопросами происхождения славян занимаются многие ученые разных специальностей: историки, археологи, языковеды, этнографы, антропологи и др. Делаются попытки восстановить этническую историю народов по предметам материальной культуры, на основании языка и письменности, по физическим типам людей.
Совокупность полученных данных приводит исследователей к разным гипотезам, которые можно разделить на две диаметрально противоположные по смыслу группы. По гипотезам одной группы, славянские народы произошли от общих предков; в соответствии с гипотезами другой группы, предки у нынешних славянских народов были разные, т.е. они не связаны общностью происхождения.
Сейчас в славянской группе народов, как, впрочем, и любых других, виден результат их длительной истории. Поэтому и не удивительно, что современные славяне антропологически неоднородны и даже принадлежат к разным ветвям европеоидной расы. Признаки, характеризующие отдельные славянские группы, проявляются и у соседних народов. Причину тому исследователи видят в расселении древних славян на обширных территориях Европы, интенсивнее всего проходившем в VI-VII вв. . В новых местах они контактировали с разными народами: с германцами на западе, с фракийцами, кельтами, иллирийцами на юге, с иранцами и финно-уграми на востоке, с балтами на севере. Естественно, что инородцы воздействовали на славян и со временем внесли собственные черты в их антропологический облик.
По археологическим и лингвистическим данным, прародиной славянских народов был Северокарпатский регион, включавший Галицию, Волынь и Подолье, откуда и началось продвижение по Европе. В VII-VIII вв., по мнению В.О.Ключевского, славяне восточной ветви, занимавшие северо-восточные склоны Карпат, стали продвигаться на восток и северо-восток Европы под натиском аваров .
Уже ко времени переселения славян в Восточную Европу соотношение исходных антропологических компонентов в их группах могло отличаться. По мере же освоения восточноевропейских территорий в разных направлениях все более проявлялся полиморфизм (разнообразие сочетаний антропологических признаков), обусловленный смешением с местным населением. Например, по комплексу расоводиагностических признаков белорусы и русские сходны с северо-западными народами (т.е. сказывается финно-угорское и летто-литовское влияние), а украинцы - с южными (из-за иранского и романского влияния) .
Становление государственности на Руси, очевидно, в немалой степени способствовало межэтническим взаимодействиям в пределах европеоидной расы. Свой вклад в метисацию внесло и освоение новых земель на востоке Евразии, где славянское население соприкасалось с монголоидным. В Восточной Европе Русь расширялась за счет колонизации северо-восточных территорий; там строились русские города и заставы, заселяемые восточными славянами "из разных мест, из разных племен". Поэтому славянское население Руси издревле различалось антропологически. Продвигаясь из Руси древней, юго-западной, в Русь новую, северо-восточную, оно смешивалось с автохтонным балтским и финно-угорским населением.
Историки полагают, что в Восточной Европе до IX в. существовали два территориальных массива славян - юго-западный и псковско-новгородский, объединение которых привело к созданию Древнерусского государства . Таким образом, исторические сведения, археологические и лингвистические данные свидетельствуют об антропологической гетерогенности исходного населения, ставшего основой русского народа. Действительно, обобщенный картографический анализ антропологических данных выявляет в составе русского населения три "ядерные структуры" - восточную, центральную и западную. Первая сформировалась на основе дославянского субстрата, вторая - за счет длительной метисации, третья - за счет исходного заселения славянским народом .
Такой, в весьма кратком изложении, вырисовывается история славянских народов, и русского в том числе, на основе археологических, лингвистических и антропологических сведений. Подтверждается она, подвергается исправлениям или вовсе отвергается результатами генетических исследований, активно проводимых в последние десятилетия? Чтобы ответить на этот вопрос, приведем здесь собственные сведения по молекулярной антропологии русского народа.
Родительские линии ДНК в генофонде русских
Этническое своеобразие любого народа обусловлено генетическими процессами, которые сиюминутно внешне себя ничем не проявляют. Но о них можно узнать, оценив сходства и отличия в детальном строении ДНК - митохондриальной (мтДНК), которая, напомним, наследуется по материнской линии, и Y-хромосомы, наследуемой по отцовской линии. Сейчас в мире уже существуют богатые базы данных, содержащие сведения о последовательности нуклеотидов в этих молекулах, на их основе строятся филогенетические древа. Относительно недавно, в последние 10-20 лет, возникла даже самостоятельная научная дисциплина - молекулярная антропология. Ее объектами и служат мтДНК и Y-хромосома, а конечной целью - генетическая история того или иного народа. Число исследований такого рода растет с каждым годом, мы тоже применяем молекулярно-генетический подход для изучения структуры геномов разных этносов, чтобы узнать их истоки .
Реконструировать процессы, которые приводят к формированию генофондов популяций, этнических групп и этнорасовых общностей, выяснить, как они распределялись пространственно за время их истории, можно с использованием больших массивов данных об изменчивости ДНК. Что касается евразийских народов, то в современный анализ филогеографической изменчивости * ДНК вовлечены много тысяч человек из различных стран и регионов. По этим вполне представительным статистическим выборкам в линии мтДНК удалось выделить монофилетические группы и подгруппы, а затем реконструировать последовательность ее эволюционных изменений.
* Филогеографическая изменчивость - это распределение вариантов ДНК этноса (популяции, группы) за время существования в определенном географическом пространстве.На сегодня уже установлено, какие митохондриальные линии присущи населению Западной и Восточной Евразии, Африки, Австралии и Америки. По современным представлениям, все евразийские группы мтДНК входят в состав трех макрогрупп - M, N и R, которые произошли примерно 65 тыс. лет назад из африканской митохондриальной группы L3. Важно, что распределение линий мтДНК в генофонде населения Евразии характеризуется выраженной этнорасовой специфичностью. Например, генофонды народов западной и восточной частей Евразии различаются кардинально. Почти все линии мтДНК корейцев и бурят относятся к набору восточноевразийских групп - A, B, C, D, F, G, M7, M8a, M9, M10, M11, Y, Z, N9a, R9. Европейцам же свойственны группы H, HV*, pre-V, T, J, K, U2, U3, U4, U5, U8, N1a, I, W и X. В другой части Евразии, в северной, судя по полученным к настоящему времени данным, лишь в краевых популяциях почти полностью преобладают митохондриальные линии одного этнорасового происхождения - монголоидного или европеоидного. Огромная же по протяженности территория (от Алтае-Саянского нагорья до Восточно-Европейской равнины) представляет собой древнюю контактную зону. Именно в ней проходил расогенез за счет межрасового смешения.
Филогенетическое дерево митохондриальной ДНК человека.
В начале 90-х годов мы занялись исследованием этнической истории русского народа, для чего изучали вариабельность материнских и отцовских линий ДНК, т.е. мтДНК и Y-хромосомы. Чтобы выявить изменчивость мтДНК в популяциях, использовали комбинированный прием: анализировали рестрикционный полиморфизм * фрагментов кодирующих районов и изменчивость нуклеотидных последовательностей некодирующих гипервариабельных участков. В результате выяснилось, что митохондриальный генофонд русского населения характеризуется высоким разнообразием, но встречались и общие группы. Наиболее частыми из них были H, U, T и J - те же группы, которые широко распространены в генофондах других европейских народов. Монголоидная примесь у русских оказалась незначительной - около 1.5%, причем ее составляли группы мтДНК (C, D, M*, G2a, N9a) восточноевразийского происхождения.
* Имеются в виду различия в наборах участков мтДНК, возникающих при ее расщеплении (рестрикции) специальными ферментами. В данном случае расщеплению подвергали не всю мтДНК, а только кодирующие участки.Необходимо было попытаться выяснить истоки разнообразия мтДНК у русских, понять, не следствие ли это особенностей формирования славянского этноса. Поэтому мы посмотрели, как распределяются гаплотипы мтДНК в европейских популяциях: русских, поляков, боснийцев, словенцев, финнов, французов, немцев. Судя по результатам этого филогеографического анализа, митохондриальные генофонды европейцев имеют общие черты. Маркеры же, которыми одни этнические группы и общности отличаются от других, представлены, как правило, комбинациями редких подгрупп и отдельных гаплотипов мтДНК. Следовательно, существует единый генетический субстрат, на основе которого развивались генофонды западных и восточных славян, а также их соседей - германцев и западнофинских народов (эстонцев и финнов) . Отдельные митохондриальные линии этого субстрата попали и в генофонд народов, населявших юг Европы и Балканы (по-видимому, в основном в процессе славянских миграций). Об этом можно судить, приняв во внимание тот факт, что южнославянские популяции отличаются генетически от их соседей - греков, албанцев и итальянцев .
Основываясь на данных об изменчивости мтДНК, мы попытались выяснить, что стало с этой молекулой при дифференциации славян на восточных, западных и южных . Кроме того, посмотрели, как сказалось такое разделение на генофондах соседних популяций Европы и Западной Азии. Оказалось, что между разными группами славян существуют не только антропологические, но и генетические различия. Их сила определяется в значительной мере степенью метисации с тем дославянским населением, которое существовало в пределах современного этнического ареала славян. Кроме того, сказывается и интенсивность, с которой они взаимодействовали с соседними народами. Последний вывод следует из того, что соседи русских, западнофинские популяции, характеризуются относительно высоким генетическим сходством с русскими, германские популяции - с западными славянами, а балканские - с южными. Проще говоря, в молекулах мтДНК соседей запечатлены характерные славянские черты - признак тесного взаимодействия пришлых и местных народов. Из всех этих результатов напрашивается вывод о единстве происхождения славян, центральное положение среди которых принадлежит западным группам. Иначе говоря, генофонд западных славян включает максимальное число типов мтДНК, встречающихся как у восточных, так и у южных славян, хотя между этими двумя группами нет высокого генетического сходства. По всей видимости, можно предположить, что генофонд западных славян ближе к исконно славянскому, но это пока всего лишь предположение.
Другая генетическая система, изменчивость которой теперь активно изучается в филогеографических исследованиях, - это нерекомбинирующая часть Y-хромосомы. С ее помощью можно составить представления о распространенности в популяциях тех или иных маркеров, которые, как и в мтДНК, несут информацию о генетической истории этнорасовых групп человека.
В своих исследованиях мы тоже анализировали изменчивость Y-хромосомы, изучая разнообразие ее линий в русских популяциях. Для этого использовали биаллельные SNP-локусы * ее ДНК, так как вариантами их полиморфизма определяется топология филогенетического дерева Y-хромосомы. Было проведено исследование около 400 индивидуальных препаратов ДНК русского населения из восьми областей - Белгородской, Орловской, Тульской, Калужской, Владимирской, Нижегородской, Ярославской и Псковской.
* SNP (Single Nucleotide Polymorphism - однонуклеотидный полиморфизм)-локусы - это участки ДНК, которые отличаются один от другого единичной нуклеотидной заменой, например: GTAC и GCAC.В результате выяснилось, что мужской генофонд русских представлен 14 группами Y-хромосомы (P*, R1*, R1a, N*, N1с, BR*, C, F*, G, I, J, H, E, K*). Однако большинство линий (80%) относятся лишь к трем из них - R1a, I и N1с. Наиболее интересными представляются группы R1a и N1с, поскольку в Европе первая маркирует славянские народы, а вторая распространена с наибольшей частотой (более 50%) у финно-угорского и балтского населения Северной и Восточной Европы и практически отсутствует в генофондах западных и южных европейцев, включая славян . Там, где славяне соседствуют с другими этническими общностями, в ДНК Y-хромосомы последних резко снижается частота встречаемости R1a . Так, например, характеризуются немцы, живущие в приграничных с поляками или с сорбами (лужицкими сорбами, или лужичанами) районах.
В генофонде русских частота встречаемости
N1с варьирует в широком диапазоне: от 5% в Калужской обл. до 35% в Псковской. Это свидетельствует об отличающейся степени взаимодействия между славянским и финно-угорским/балтским населением в разных частях восточноевропейского ареала русских. Из проанализированных нами образцов Y-хромосомы лишь в псковской и поморской (этнографической группы русского населения Архангельской обл.) популяциях частота гаплогруппы
N1с оказалась сопоставимой с той, что характеризует финно-угров и балтов .
Примечательно, что более тонкими исследованиями полиморфизма Y-хромосомы можно различить две группы в N1с - "балтскую" и "финно-угорскую" . Нам удалось это сделать по результатам анализа 12 STR-локусов *, которые эволюционируют с более высокой скоростью, чем упомянутые биаллельные SNP-локусы. Оказалось, что высокая частота гаплогруппы
N1с в псковской популяции обусловлена преимущественно "балтскими" вариантами Y-хромосомы (их доля составляет 64%). В генофондах русского населения центральных и южных областей европейской части России преобладают (в среднем до 80%) N1с -варианты Y-хромосомы, характерные для финно-угорского населения Восточной Европы.
* STR-(Short Tandem Repeats - короткие тандемные повторы)-локусы - это относительно короткие (до 6 пар нуклеотидов) участки ДНК, расположенные в молекуле в виде блоков из повторяющихся последовательностей. Иначе STR-локусы называют микросателлитными последовательностями ДНК.Итак, у славянских народов набор маркеров в ДНК Y-хромосомы сходен. Но, что важно, существующие отличия славян от соседей (германцев на западе Европы; греков, албанцев и итальянцев на юге; финно-угорского населения на востоке Европы) по Y-хромосоме гораздо заметнее, чем по линиям мтДНК .
Дифференциация родительских линий ДНК
Наиболее информативен для исследований судьбы этносов анализ генетической дифференциации популяций по данным об изменчивости мтДНК и Y-хромосомы. Результаты такого рода анализа позволяют составить представление о том, как распространялись материнские и отцовские линии ДНК в процессе экспансии народов. Хороший пример в этом отношении - русские, освоившие за последние почти полторы тысячи лет обширные пространства Евразии.
Мы изучили мтДНК и Y-хромосомы в 10 популяциях русского населения Восточной Европы: от Ставропольского края на юге до Псковской обл. на севере и от Орловской обл. на западе до Нижегородской на востоке. Кроме собственных результатов, мы привлекли в анализ данные о поморской, костромской, ростовской и курской популяциям, полученные другими научными коллективами.
В итоге выяснилось, что по отцовским линиям русское население отличается в значительно большей степени, чем по материнским. Так, коэффициенты дифференциации FST * Y-хромосомы составляют 3.1%, в то время как мтДНК - лишь 0.42%, т.е. различаются почти на порядок. Примечательно в этих коэффициентах еще одно: по Y-хромосоме у русских они существенно ниже, чем в Европе в целом (там их величина достигает 7.04%), а по митохондриальным линиям не отличаются (0.42% и 0.41%).
* FST - это коэффициент локальной генетической дифференциации, мера генетической подразделенности популяции и одновременно эквивалент инбридинга субпопуляции (S) относительно всей популяции (T).Хотя в популяциях русского населения различия в распределении материнских линий ДНК не очень высоки, статистическим анализом нам удалось выявить две ветви - юго-западную и северо-восточную. Каждая из них образует кластер с другими европейскими популяциями. Так, юго-западная ветвь мтДНК входит в кластер материнских линий, свойственных народам Центральной Европы, - как западнославянским (например, полякам), так и балтским (литовцам), а также некоторым финно-угорским (эстонцам). Северо-восточная ветвь мтДНК русских проявляет сходство с мтДНК финно-угорских и других народов Восточной Европы (финнов, карелов, марийцев, татар, адыгейцев).
Результаты анализа FST-дистаций, полученных на основании полиморфизма биаллельных локусов Y-хромосомы, также свидетельствуют о гетерогенности русских популяций. Однако картины дифференциации по мтДНК и Y-хромосоме не совпадают. В сравнительном анализе использовались литературные данные ] об изменчивости Y-хромосомы украинцев, белорусов, литовцев, латышей, эстонцев, финнов, марийцев и чувашей, т.е. соседних с русскими этнических групп. Собственные сведения о полиморфизме Y-хромосомы русского населения в Восточной Европе мы дополнили также данными об изменчивости этой хромосомы у поморов . Из анализа всей совокупности ее характеристик следует, что лишь псковская и поморская популяции проявляют выраженное сходство с финно-угорскими и балтскими народами Северной и Восточной Европы. Подавляющее же большинство русских популяций по особенностям генетических структур образуют единый кластер вместе с такими славянскими народами, как поляки, украинцы и белорусы. У всех у них высока частота группы R1a, а группа N1с встречается с умеренной или даже низкой частотой. Эти же группы Y-хромосомы в псковской и поморской популяциях встречаются в обратной пропорции (как у финно-угорских и балтских народов): частота
N1с повышена, а R1a - понижена.
Структура митохондриального генофонда русских и еще двух славянских народов - поляков и боснийцев.
Кластеры (группы и подгруппы) мтДНК обозначены буквами;
розовыми кружками (их размер пропорционален частоте гаплотипа мтДНК) даны кластеры
русских, серыми - поляков, черными - боснийцев.
Кластеры (группы и подгруппы) мтДНК обозначены буквами;
розовыми кружками (их размер пропорционален частоте гаплотипа мтДНК) даны кластеры
русских, серыми - поляков, черными - боснийцев.
Таким образом, результаты комбинированного анализа позволяют заключить, что в русских популяциях центральноевропейский генетический компонент как материнского, так и отцовского происхождения выражен по-разному. Этот компонент преобладает лишь в южной и западной частях восточноевропейского ареала русских. В центре и на востоке ареала начинают доминировать восточноевропейские линии мтДНК. И только в некоторых популяциях северной части ареала русских комбинация линий мтДНК и Y-хромосомы та же, что у финно-угорского населения. Выявленная картина генетической дифференциации современных русских, по-видимому, отражает процессы, обусловленные межэтническим взаимодействием в ходе формирования русского народа начиная с эпохи средневековья.
Итак, мы выяснили молекулярно-генетическую структуру русского генофонда, правда, пока лишь в считанном количестве популяций. В целом она согласуется с результатами картографического анализа антропологических данных, ставшими основой гипотезы об истории формирования русского народа. Судя по всему, лишь на западе этнической территории русские представляют собой генетических потомков славян, в центральной части русский этнос сформировался за счет смешения славян и дославянского финно-угорского населения, а на востоке и в северной части ареала произошло только замещение дославянских языков и культуры славянскими . Между тем молекулярно-генетические данные позволили глубже понять причины наблюдаемых генетических и антропологических различий. Исходя из распределения линий мтДНК и Y-хромосомы в русских популяциях, становится очевидным, что наблюдаемая "трехзональная" картина их дифференциации обусловлена различным участием мужчин и женщин в освоении восточноевропейских территорий. По всей видимости, лишь начальный этап колонизации осуществлялся как мужчинами, так и женщинами (видимо, племенными группами, родами, семьями), а более поздние - преимущественно мужчинами, которые женились на аборигенках.
Расположение европейских популяций в пространстве двух координат.
"Карты" построены на основании результатов многомерного шкалирования (одного из методов многомерной статистики) межпопуляционных FST-дистанций по данным об изменчивости мтДНК (вверху) и Y-хромосомы. Для анализа использованы также литературные данные . Видно, что по мтДНК русские, живущие в разных областях, не очень значительно отличаются друг от друга, но все же разделяются на юго-западную и северо-восточную зоны. В первой, кроме русских, оказываются некоторые народы из западных славян (поляки), балтов (литовцы) и финно-угров (эстонцы). Во вторую зону вместе с русскими популяциями попадают несколько финно-угорских народов, а также татары и адыгейцы. Расположение популяций по Y-хромосоме отражает своеобразие их мужского генофонда, определяемого, главным образом, высокой частотой группы N1с (61%) и очень низкой (10%) - R1a. Если сравнить обе "карты", становятся очевидными генетические различия между генофондами популяций по мтДНК и Y-хромосоме. Особенно это касается финнов: в первом случае они располагаются среди русских Ярославской, Владимирской и Тульской областей, а по Y-хромосоме занимают обособленное положение .* * *Полученная в нашей работе картина генетической дифференциации русского населения Восточной Европы нуждается, конечно, в уточнениях. В связи с этим нужно изучить в дальнейшем изменчивость мтДНК и Y-хромосомы в тех русских популяциях, которые еще не охвачены такими исследованиями. Кроме того, необходимо узнать, каким было генетическое разнообразие древнего восточноевропейского населения в разные периоды его истории, начиная от дославянского.
Как показывают результаты анализа ДНК, генетическая преемственность между древними и современными народами соблюдается отнюдь не всегда, ведь одни народы могли сменяться другими. Например, генофонды неолитического и нынешнего населения Прибайкалья имеют мало общего . На возможную смену этносов в некоторых европейских регионах - Тоскании (Италия) и Стране басков (Испания) - указывают результаты генетических исследований древнего и современного населения . Более того, уже имеются примеры существенных противоречий между наблюдаемыми и ожидаемыми распределениями маркеров мтДНК в палеопопуляциях. Так, у неолитического населения Страны басков не обнаружена митохондриальная группа V. А ведь именно ей придается большое значение, когда, основываясь на распределении групп мтДНК у современных европейцев, моделируют процесс послеледникового заселения Европы выходцами из пиренейского "убежища" .
Выявлено и еще одно серьезное несоответствие. У неолитического населения Центральной Европы, ведущего происхождение от "первых европейских фермеров" (носителей переднеазиатской технологии агрокультуры), митохондриальная группа N1a встречается с неожиданно высокой частотой. Современным же европейцам эта группа практически не свойственна и потому никак нельзя было предсказать ее высокую распространенность среди неолитического населения .
Приведенные примеры показывают, насколько важна проверка этноисторических моделей, если выявленное генетическое разнообразие современных народов переносится в их прошлое. Выходит, экстраполяции не всегда надежны. Каким был генофонд далеких предков, безошибочно можно судить по результатам исследований ДНК непосредственно древнего населения. И такой анализ вполне возможен, ведь археологами найдено множество останков неолитического времени, пригодных для выделения ДНК.
Из работы ХРОМОВОЙ НАТАЛЬИ АНАТОЛЬЕВНЫ;
ПОЛИМОРФИЗМ СИСТЕМЫ HLA У ПРЕДСТАВИТЕЛЕЙ РАЗНЫХ СЛАВЯНСКИХ ЭТНИЧЕСКИХ ГРУПП (РУССКОЙ, БЕЛОРУССКОЙ И УКРАИНСКОЙ)
Комплекс генов HLA (главного комплекса гистосовместимости человека) компактно расположен на коротком плече 6-й аутосомной хромосомы, занимает 3500 kb (тысяч пар оснований) и содержит более 220 генов . В состав HLA входят три группы генов: экспрессирующиеся гены, псевдогены и гены с неустановленной функцией . Гены локуса HLA -DP, -DQ и –DR, ближайшие к центромере, кодируют молекулы II класса. Молекулы II класса определяются на поверхности так называемых антиген-представляющих (вспомогательных) клеток - дендритных, активированных макрофагов, В-лимфоцитов, а также активированных эндотелиальных, эпителиальных и тучных клеток, Т-хелперов .
Система HLA является одной из наиболее полиморфных генетических систем, выполняющей в организме человека ряд функций, важнейшими из которых являются генетический контроль иммунного ответа и поддержание иммунного гомеостаза, нарушение которого лежит в основе таких патологических процессов, как аутоиммунные заболевания и развитие опухолей . Обеспечивая регуляцию иммунного ответа, гены HLA осуществляют генетический контроль взаимодействия всех иммунокомпетентных клеток организма, распознавание своих и чужеродных (в том числе измененных собственных) клеток, запуск и реализацию иммунного ответа и, в целом, обеспечивают выживание человека как вида в условиях экзогенной и эндогенной агрессии . Эти функции, в первую очередь, обусловлены двумя свойствами системы HLA: участием в презентации антигенного пептида и широким разнообразием – полиморфизмом аллельных вариантов каждого гена.
Для «запуска» адаптивного иммунного ответа на патоген, его антигенный пептид для распознавания Т-хелпером должен быть представлен антиген представляющей клеткой только в контексте с молекулой HLA II класса .
Несмотря на мощное действие направленного отбора, полиморфизм системы HLA сохраняется и является необходимым элементом при взаимодействии человека с окружающей средой. Каждый из аллелей генов HLA обеспечивает возможность реагировать на определенный набор пептидов антигенов, и индивидуумы, которые гетерозиготны по HLA, могут иметь эффективный иммунный ответ на более разнообразный спектр антигенов и имеют намного больше шансов противостоять инфекции, что способствует поддержанию HLA-разнообразия на высоком уровне.
Исследование полиморфизма системы HLA является одним из наиболее эффективных подходов к изучению структуры и функции главного комплекса гистосовместимости человека. Необходимость подобных исследований обусловлена, в том числе и тем фактом, что без полного представления о строении системы генов тканевой совместимости невозможно успешное развитие клинической трансплантологии, и в первую очередь, это касается пересадки аллогенного костного мозга.
Для изучения полиморфизма системы HLA класса II используется, в основном, анализ особенностей распределения DRB1 гена. Объяснением этого факта может служить значительно более выраженный полиморфизм гена DRB1, по сравнению с DQA1 и DQB1 генами. Гаплотипичесие сочетания DRB1-DQA1-DQB1, вследствие их еще большей вариабельности, лучше отражают разнообразие HLA-системы.
В последние 25 лет появилась возможность исследования HLA на качественно новом молекулярно-генетическом уровне при помощи полимеразной цепной реакции. Принципиальным отличием новых методов явилось использование в качестве объекта исследования непосредственно генетического материала (участков ДНК, определяющих аллельный полиморфизм системы HLA) . Результатом этого стало заметное увеличение количества известных специфичностей HLA - со 150 в 1991 году , 472 в 1998 году до примерно 2500 (данные на 2006 год) аллелей . При этом среди вновь открытых аллелей установлены аллели чрезвычайно выраженной ассоциации с заболеваниями.
Исследования полиморфизма HLA в различных этнических группах позволяют обнаруживать все новые аллельные варианты генов HLA. В последние годы благодаря распространению в России молекулярно–генетических методов изучения HLA появилась возможность исследования аллельного разнообразия генов HLA в разных этнических группах, в том числе славянских, проживающих как на территории России, так и вне ее.
Данные, полученные в результате изучения распределения генов II класса главного комплекса гистосовместимости в различных этнических группах, могут быть использованы для развития клинической трансплантологии, направления “HLA и болезни”, криминалистики (идентификация личности) и такой фундаментальной науки, как антропология, поэтому представлялось целесообразным проведение исследования, посвященного изучению HLA–разнообразия в популяциях, принадлежащих к различным этническим группам, на современном молекулярно–генетическом уровне.
Изучая полиморфизм специфичностей генов HLA II класса (DRB1, DQA1 и DQB1) и их гаплотипических сочетаний в трёх различных восточнославянских этнографических популяциях: русской, белоруской и украинской с помощью ранее созданных в ГНЦ «Институт иммунологии ФМБА России» вариантов ДНК-типирования (выделение геномной ДНК из периферической крови, типирование генов HLA класса II методом мультипраймерной полимеразной цепной реакции, электрофорез продуктов ПЦР в агарозном геле) с целью последующего использования полученных данных для развития фундаментальной биологической науки (антропология и иммуногенетика) и для прикладной медицины (клиническая трансплантология, «HLA и болезни»).
Задачи исследования:
1. Провести анализ распределения специфичностей HLA- DRB1, DQA1 и DQB1, а также их гаплотических сочетаний у здоровых представителей русской, белоруской и украинской популяций в сравнении с аналогичными данными для других популяционных групп.2. Проанализировать генетическое родство исследованных популяций, как между собой, так и с некоторыми народами Европы.
3. На примере восточных славян изучить способность системы генов HLA отражать геногеографию этнических групп и возможность использования этой системы в популяционных исследованиях.
В процессе работы впервые был проведен сравнительный анализ особенностей распределения специфичностей генов HLA II класса (DRB1, DQA1 и DQB1) и их гаплотипических сочетаний на молекулярно–генетическом уровне среди представителей белорусских, украинских и русской популяций.
На основе полученных данных был проведен расчет генетических расстояний, что позволило оценить генетическое родство между исследованными этническими группами:
Оценено генетическое родство (по профилю распределения гена DRB1) исследованных популяций с некоторыми народами Европы и выявлены межпопуляционные различия.
Впервые на примере восточных славян, показано, что, несмотря на влияние направленного естественного отбора, распределение генов HLA II класса адекватно отражает геногеографию этнических групп, и особенности полиморфизма генов HLA могут быть использованы в популяционных исследованиях.
Впервые получены данные по распределению генов HLA II класса для восточнославянских популяций, которые могут быть использованы в качестве контрольных для поиска маркеров генетической предрасположенности к развитию различных заболеваний, для идентификации личности.
Полученные данные могут быть использованы для изучения истории формирования таких генетически близких народов, как восточные славяне.
МАТЕРИАЛЫ И МЕТОДЫ ИССЛЕДОВАНИЯ
Для исследования были использованы образцы крови взрослых здоровых и неродственных между собой представителей пяти популяций, представляющих три восточнославянских этноса: две популяции белорусов (Витебская и Брестская области), две популяции украинцев (Львовская и Хмельницкая области) и русская популяция (Вологодская область).
Места обследования планировались так, чтобы изученные популяции дали представление о важнейших подразделениях генофонда каждого народа. Среди белорусов антропологи выделяют две важнейших группы – северных (представленных у нас выборкой из Витебской области) и южных (представленных у нас выборкой из Брестской области). Брестская популяция представляет белорусское Полесье. Антропологическое подразделение украинского этноса, в отличие от белорусского следует не по оси «север-юг», а по оси «запад-восток». В соответствии с этим нами были обследованы популяции, представляющие западный вариант антропологического типа украинцев (Львовская область) и восточный вариант (Хмельницкая область). Русская популяция представлена населением Вологодской области, которое, относясь в целом к северному варианту русского генофонда, в течение многих веков впитывало в себя и генные потоки из среднерусской полосы.
Каждая из пяти популяций представлена только коренным сельским населением, относящимся к целому ряду населенных пунктов и потому репрезентативно представляющим данную часть этнического ареала. В выборку включались только те неродственные между собой индивиды, все бабушки и дедушки которых относятся к данному этносу и родились в пределах данной популяции.
Особенности распределения специфичностей гена DRB1 в исследованных популяциях.
Данные распределения специфичностей гена DRB1 представлены в табл. 1
Таблица 1. Распределение специфичностей гена DRB1 в исследованных популяциях.
Специфичности
DRB1
|
Белор. сев. (Витебс-кая область),
N=100 (%)
|
Белор. южн. (Брестс-кая область),
N=105 (%)
|
Укр. вост. (Хмельниц-кая область)
N=137 (%)
|
Укр. запад. (Львовс-кая область),
N=102 (%)
|
Русские (Вологод-ская область),
N=121 (%)
|
*01
|
10,0
|
10,5
|
9,5
|
12,2
|
12,4
|
*03
|
7,0
|
9,0
|
5,5
|
7,8
|
7,4
|
*04
|
7,5
|
12,9
|
10,9
|
11,3
|
14,0
|
*07
|
10,5
|
10,5
|
14,6
|
15,2
|
14,9
|
*08
|
5,5
|
2,9
|
3,6
|
4,4
|
5,4
|
*09
|
0,0
|
1,9
|
0,4
|
1,5
|
2,5
|
*10
|
0,0
|
0,5
|
1,1
|
0,0
|
0,4
|
*11
|
15,0
|
13,3
|
18,6
|
18,6
|
9,5
|
*12
|
4,0
|
3,8
|
2,2
|
0,5
|
1,2
|
*13
|
16,5
|
11,9
|
12,4
|
10,8
|
14,9
|
*14
|
1,0
|
2,9
|
2,2
|
2,0
|
1,2
|
*15
|
18,0
|
13,3
|
10,2
|
9,8
|
14,5
|
*16
|
5,0
|
6,7
|
8,8
|
5,9
|
1,6
|
При анализе частотного распределения аллелей гена DRB1 среди представителей северной белорусской популяции (Витебская область) выявлено, что самыми частыми аллелями являются DRB1*15 (18,0%), DRB1*13 (16,5%) и DRB1*11 (15,0%), относительно распространенными являются DRB1*07(10,5%), DRB1*01(10,0%), редко встречается DRB1*14(1,0%), не определяются DRB1*09 и DRB1*10. В популяции белорусского Полесья (Брестская область) наиболее частыми представлены следующие аллели - DRB1*15 и DRB1*11 (с частотой 13,3% соответственно), DRB1*13 (11,9%), также отмечается высокое значение DRB1*04 (12,9%). Относительно распространенными являются DRB1*07 и DRB1*01 (10,5% соответственно), редко встречающимися - DRB1*09(1,9%), DRB1*10(0,5%) и DRB1*14(2,9%).
Среди представителей восточного варианта украинской популяции (Хмельницкая область) наиболее распространенными аллелями гена DRB1 являются DRB1*07(14,6%), DRB1*11(18,6%) и DRB1*13(12,4%), относительно распространенными - DRB1*04(10,9%) и DRB1*15(10,2%), менее распространенными - DRB1*09(0,4%), DRB1*10(1,1%), DRB1*12 и DRB1*14 (с частотой 2,2% соответственно). В западном варианте украинской популяции (Львовская область) часто встречаемыми отмечаются аллели DRB1*01(12,2%), DRB1*04(11,3%), DRB1*07(15,2%) и DRB1*11(18,6%), редко встречаемыми - DRB1*09(1,5%), DRB1*12(0,5%) и DRB1*14(2,0%). Остальные аллели по частоте встречаемости занимают промежуточное положение, DRB1*10 не определяется.
Наиболее распространенными аллелями гена DRB1 среди представителей русской популяции (Вологодская область) отмечаются следующие особенности. Наиболее распространенными аллельными вариантами являются DRB1*04(14,0%), DRB1*07(14,9%), DRB1*13(14,9%) и DRB1*15(14,5%), редко встречающимися - DRB1*10(0,4%), DRB1*12(1,2%), DRB1*14(1,2%) и DRB1*16(1,6%). Остальные аллели по частотам имеют промежуточные значения.
Особенности распределения специфичностей гена DQA1 в исследованных популяциях.
При анализе частотного распределения аллельных вариантов локуса DQA1 среди представителей белорусских популяций выявлены следующие особенности. Наиболее частыми аллелями в северной популяции (Витебская область) являются DQA1*102(28,5%) и DQA1*501(26,5%), редко встречающимися - DQA1*601(2,0%), остальные аллели по частоте встречаемости занимают промежуточные значения. Среди представителей южной белорусской популяции (Брестская область) отмечаются широко распространенные аллели DQA1*501(29,5%), DQA1*102(19,5%) и DQA1*301(14,8%), редко встречаемые - DQA1*401(2,9%) и неопределяемые - DQA1*601.
При анализе особенностей распределения аллельных вариантов гена DQA1 среди представителей украинских популяций выявлено, что самыми распространенными являются DQA1*0501, DQA1*0102, DQA1*0201 (28,8%, 20,1% и 14,6% соответственно для восточного варианта украинской популяции; 30,4%, 17,7% и 15,2% соответственно для западного варианта). Редко встречается аллель DQA1*0401 (с частотой 2,6% и 2,0% для каждой группы), аллель DQA1*0601 отсутствует в обеих популяциях.
Самыми распространенными аллелями локуса DQA1 у русских являются DQA1*0501(21,1%), DQA1*0102(18,6%) и DQA1*0301(16,9%). Остальные по частотам аллели имеют средние значения, частота аллеля DQA1*0601 имеет наименьшее значение.
Особенности распределения специфичностей гена DQB1 в исследованных популяциях.
В локусе DQВ1 в северной белорусской популяции наиболее распространенными выявлены DQВ1*0602-8(31,5%), DQВ1*0301(23,0%) и DQВ1*0201(14,5%), редко распространенными - DQВ1*0304(0,5%), DQВ1*0503(1,0%) и DQВ1*0601(1,5%); DQВ1*0305 не определялся. В популяции белорусского Полесья часто встречаемыми аллелями также являются DQВ1*0602-8(20,0%), DQВ1*0301(22,4%) и DQВ1*0201(17,1%), редко встречаемыми - DQВ1*0304(0,5%), DQВ1*0503(2,9%) и DQВ1*0601(1,9%); DQВ1*0305 также не определялся.
В локусе DQВ1 в восточном варианте украинской популяции (Хмельницкая область) наиболее распространенными аллелями являются DQВ1*0201(18,3%), DQВ1*0301(26,3%) и DQВ1*0602-8(19,0%), менее распространенными - DQВ1*0303(2,6%), DQВ1*0503(2,2%) и DQВ1*0601(0,7%); в западном варианте (Львовская область) самыми распространенными также являются DQВ1*0201(19,6%), DQВ1*0301(23,5%) и DQВ1*0602-8(16,7%); менее распространенными - DQВ1*0401/2(2,0%), DQВ1*0503(1,0%) и DQВ1*0601(0,5%). Аллели DQВ1*0304 и DQВ1*0305 не определяются в обеих популяциях.
В локусе DQB1 в русской популяционной группе часто встречаемые аллели представлены DQB1*0201(17,8%), DQB1*0301(16,1%) и DQB1*0602-8(25,6%), остальные аллели по частотам встречаемости занимают промежуточные положения. Частоты аллелей DQB1*0305, DQB1*0601 имеют наименьшие значения (0,8% и 0,4% соответственно), DQB1*0304 отсутствует в популяции.
При общем анализе распределения аллельных вариантов DRB1 локуса во всех представленных восточнославянских популяциях можно отметить следующие закономерности. Наблюдается высокие частоты для некоторых аллельных вариантов: DRB1*01, DRB1*07, DRB1*11, DRB1*13 и DRB1*15, что соответствует типичному западно-евразийскому варианту распределения . Частота встречаемости аллеля DRB1*04 во всех популяциях колеблется от 8,0% до 14,0%, что соответствует средним значениям в большинстве популяций мира . Обращает на себя внимание отсутствие или низкие частоты DRB1*09 и DRB1*12, которые характерны исключительно для монголоидных популяций [Lee J., 1997; Han H. et al., 1997]. В локусе DQA1 для всех исследованных популяций характерны высокие значения встречаемости следующих аллелей: DQA1*0102, DQA1*0201, DQA1*0501. При изучении распределения аллелей гена DQB1 обнаружено, что самыми частыми для всех исследованных популяций (белорусских, украинских и русских) являются DQB1*0201, DQB1*0301 и DQB1*0602-8.
Сравнительный анализ данных типирования по трем генам HLA – DRB1, DQA1 и DQB1 восточнославянских популяций показал, что основные различия между ними связаны с геном DRB1 и в меньшей степени с DQA1 и DQB1. Объяснением этого факта может служить значительно более выраженный полиморфизм гена DRB1 по сравнению с генами DQB1 и особенно DQA1. Вследствие малой информативности данных, полученных в результате типирования генов DQA1 и DQB1, сравнительный анализ частот их специфичностей не представлен в работе.
При попарном сравнении (0,05 и 0,01 уровни значимости) распределения специфичностей гена DRB1 между исследованными восточнославянскими популяциями было установлено, что достоверно различаются восточные украинцы из Хмельницкой области и русские из Вологодской области частотным содержанием DRB1*11 и DRB1*16, западные украинцы из Львовской области и русские из Вологодской области - частотным содержанием DRB1*11
Особенности полиморфизма генов HLA II класса легли в основу расчета генетических расстояний между изученными популяциями. Расчет генетических расстояний по частотам аллелей DRB1 локуса был проведен по методу М. Нея . Дендрограмма, построенная на основе генетических расстояний, представлена на рис. 1
Результаты многомерного статистического анализа выявили, что изученные нами популяции образуют два кластера, соответствующие географической и этногенетической близости популяций. «Южный» кластер включил в себя обе украинские популяции. «Северный» кластер включил обе белорусские (Витебской и Брестской областей) и русскую (Вологодской области) популяции. Неожиданной оказалась несколько большая близость русских Вологодской области к популяции белорусского Полесья, а не к северным белорусам. Сходство популяции русских из Вологодской области с белорусами Полесья может быть связано с существенным генетическим вкладом «южной» колонизации Вологодских земель по сравнению с «северной». А положение на графике северных белорусов может быть связано с влиянием балтских племен, фиксируемые антропологами на севере Белоруссии.
Рис.1 Дендрограмма генетических расстояний на основании частот аллелей DRB1 локуса у представителей восточнославянских популяций
Особенности распределения трехлокусных гаплотипов в исследованных популяциях.
В выборках русских, украинцев и белорусов были определены частоты трехлокусных гаплотипов (DRB1 - DQA1 - DQB1)
Сравнительный анализ представленных данных выявляет наличие общих и индивидуальных особенностей распределения гаплотипов в каждой популяции.
Среди белорусских популяций определяется высокая частота гаплотипов: DRB1*11-DQA1*0501-DQB1*0301 и DRB1*15-DQA1*0102-DQB1*0602-8, а также DRB1*01-DQA1*0101-DQB1*0501.
В украинских популяциях отмечается высокие частоты гаплотипов DRB1*11-DQA1*0501-DQB1*0301 и DRB1*01-DQA1*0101-DQB1*0501, а также гаплотипа DRB1*07-DQA1*0201-DQB1*0201, а среди западных украинцев (Львовская область) выявлена высокая частота гаплотипа DRB1*04-DQA1*0301-DQB1*0302.
В русской популяции обнаружены высокие частоты гаплотипов: DRB1*01-DQA1*0101-DQB1*0501, DRB1*15-DQA1*0102-DQB1*0602-8, а также DRB1*04-DQA1*0301-DQB1*0302 и DRB1*07-DQA1*0201-DQB1*0201.
В среднем наблюдается однотипность частотного распределения трехлокусных гаплотипов DRB1-DQA1-DQB1 у всех представленных популяций. Отличительной особенностью является отсутствие или низкие частоты гаплотипов: DRB1*09-DQA1*0301-DQB1*0303 и DRB1*12-DQA1*0501-DQB1*0301, которые более характерны для монголоидной популяции.
Несмотря на определенную схожесть частотного распределения, в отдельных популяциях отмечается наличие гаплотипов, которые отсутствуют у других, например: в северной белорусской популяции (Витебская область) - DRB1*12-DQA1*0601-DQB1*0301, в полесской популяции (Брестская область) - DRB1*01-DQA1*0102-DQB1*0502/4, DRB1*03-DQA1*0501-DQB1*0301, в восточной украинской популяции (Хмельницкая область) - DRB1*11-DQA1*0103-DQB1*0602-8, DRB1*15-DQA1*0102-DQB1*0502/4, у западных украинцев (Львовская область) - DRB1*09-DQA1*0301-DQB1*0201, DRB1*14-DQA1*0101-DQB1*0501 и DRB1*14-DQA1*0101-DQB1*0502/4, в северной русской популяции (Вологодская область) - DRB1*04-DQA1*0301-DQB1*0305.
Сравнение частоты трехлокусных гаплотипов в исследованных популяциях.
При попарном сравнении (0,05 и 0,01 уровни значимости) распределения трехлокусных гаплотипов (DRB1-DQA1-DQB1) между исследованными восточнославянскими популяциями было установлено, что достоверно различаются восточные украинцы из Хмельницкой области и русские из Вологодской области частотным содержанием следующих гаплотипов: DRB1*11-DQA1*0501-DQB1*0301 и DRB1*16-DQA1*0102-DQB1*0502/4.
Изучение распределения гаплотипических сочетаний показало, что наиболее информативным был сравнительный анализ трехлокусных гаплотипов. Исходя из этого, было бы целесообразным провести сравнение распределения трехлокусных гаплотипических сочетаний в исследованных популяциях с аналогичными данными из литературных источников. Однако данные по трехлокусным гаплотипам в доступной нам литературе практически не были представлены, поэтому в анализ были включены сведения о HLA–профиле некоторых популяций по локусу DRB1 как наиболее полиморфному из исследованных.
Результаты многомерного кластерного анализа изученных популяций в сравнении с некоторыми народами Европы выявили следующие особенности. Все популяции, представленные на дендрограмме (рис.2), образовали три кластера: «североевропейский», «центрально-европейский» и «южно-европейский». Русские из Вологодской области вошли в «североевропейский» кластер, наряду с немцами, датчанами, голландцами, финнами, ирландцами и англичанами. «Центрально-европейский» кластер включил в себя белорусов, чехов, бельгийцев, австрийцев, швейцарцев, французов, хорватов, украинцев, словаков и венгров. Внутри этого кластера в зависимости от генетической близости популяции распределяются следующим образом: белорусы (Витебская и Брестская области) составляют одну группу с чехами, бельгийцами, австрийцами и швейцарцами, украинцы (Хмельницкая и Львовская области) - со словаками и венграми. В «южно-европейский» кластер вошли болгары, греки и итальянцы.
Те же данные представлены в виде двухмерного графика многомерного шкалирования. (рис.3) Центральное место на графике занимают популяции, составляющие «центрально-европейский» кластер. Северные белорусы (Витебская область) несколько отдалены от основного «ядра» популяций, что, вероятно, обусловлено своеобразием указанной популяционной группы (возможным влиянием балтских племен) или недостаточным размером выборки, взятой для анализа.
С одной стороны к «центрально-европейскому» кластеру примыкают «североевропейские» популяции, из которых ближе всех к центру расположены русские из Вологодской области. Положение русских на графике свидетельствует о сочетании в генофонде указанной группы «североевропейских» и «среднеевропейских» генов. С другой стороны от «ядра» основных популяций расположились «южно-европейские» популяции: болгары, греки и итальянцы.
Подобную картину можно объяснить особенностями расселения и освоения земель древними славянами и историей формирования изученных популяционных групп.

Рис.2 Дендрограмма генетических расстояний на основании частот аллелей DRB1 локуса у представителей восточнославянских популяций и некоторых народов Европы
Рис 3. График многомерного шкалирования генетических расстояний на основе частот гена DRB1 у представителей восточнославянских популяций и некоторых народов Европы.
Бел. сев. – белорусы северные (Витебская область), Бел. южн. – белорусы Полесья (Брестская область), Укр. вост. – украинцы восточные (Хмельницкая область), Укр. зап. – украинцы западные (Львовская область), Рус. – русские (Вологодская область), Болг. - болгары, Хорв. – хорваты, Слов. – словаки, Венг. - венгры, Бельг. – бельгийцы, Австр. – австрийцы, Нем. – немцы, Франц. – французы, Датч. – датчане, Гол. – голландцы, Фин. – финны, Швейц. – швейцарцы, Ирлан. – ирландцы, Англ. – англичане, Итал. - итальянцы
Представленный в этой работе анализ полиморфизма аллелей генов HLA можно рассматривать как часть популяционных исследований, проводимых среди восточнославянских этносов: антропологические исследования, анализ митохондриальных и Y-хромосомных, а также других аутосомных маркеров ДНК. Наиболее полная информация по восточнославянским народам была получена в ходе нескольких антропологических экспедиций: «Русской антропологической экспедиции» (В.В. Бунак, 1965), «Украинской и Прибалтийской антропологических экспедиций» (Дьяченко, 1965, Витов, Чебоксаров, 1959). Для исследований были использованы следующие методы: оценка антропологических признаков (размеры головы и лица, показатели цвета волос и глаз, интенсивности роста бороды, форма носа), дерматоглифика, география фамилий и т.п. Ареал славянских народов находится в рамках той обширной зоны, которая разделяет ареалы северных и южных европеоидов, выделяющихся, в пределах европеоидной расы, максимально светлой и максимально темной пигментацией. Население этой переходной зоны отличается большим разнообразием локальных сочетаний антропологических признаков.
Принято объединять эти локальные варианты в две группы – западноевропейскую и центрально-восточноевропейскую. На территории Восточной Европы распространены, в основном, четыре расы: среднеевропейская, альпо-карпатская, индо-средиземноморская и лапоноидная. Каждая из них, за исключением лапоноидной, представителями которой являются саамы, представлена рядом антропологических типов. На крайнем востоке Европы у многих из этих типов проявляется древняя уралоидная примесь, которая может сочетаться с понтийским компонентом (характерным для южных европеоидов), а также - с относительно недавним влиянием южно-сибирской монголоидности.
На северо-западе Восточной Европы по побережью Балтики среднеевропейская раса включает атланто-балтийский компонент. В беломоро-балтийской зоне черты северных европеоидов сочетаются с определенным уралоидным компонентом. У восточнославянских народов наиболее заметно присутствуют черты среднеевропейской расы. На юго-западе расселения восточных славян присутствует заметное участие альпо-карпатского компонента. На юге и юго-востоке – понтийского, который может быть отмечен у болгар и албанцев, живущих на Украине; на востоке – субуральского, на севере – беломоро-балтийского [Алексеева Т.И., 1999].
В настоящее время широко используется относительно новый метод - геногеография, где для анализа используются генетически наследуемые мало полиморфные системы, в частности система ABO, некоторые биохимические маркеры. Информация о генофонде населения Восточной Европы содержит данные из литературных источников, опубликованных с 1985 по 1995 гг. Геногеография народонаселения способна напрямую связывать генетическую действительность современных поколений с древностью. В результате исследования было обнаружено, что из всех воздействий на генофонд за все время существования населения Восточной Европы главное формообразующее значение имели воздействия в направлении запад-восток. Они носили характер проникновения с запада – юго-запада и распространения по Восточной Европе генных комплексов европейского и балканского происхождения и их носителей – древних народов и племен. Население Восточной Европы постепенно и достаточно равномерно генетически становилось европейским, но эта генетическая «европеизация» не дошла до конца. Возникло некое равновесие между европейскими и азиатскими влияниями, возникло своего рода евразийское равновесное состояние генофонда народов Восточной Европы [Алексеева Т.И., 1999].
Несмотря на то, что типирование генов HLA – относительно новый подход, создающий новые возможности в оценке генетического профиля популяций, трудно сравнивать результаты, представленные в этой работе, с данными анализа других генетически наследуемых факторов, полученными в ходе многолетних исследований из-за разницы в количестве обследованных популяционных групп. Тем не менее, даже такое небольшое количество изученных популяций отражает основные закономерности геногеографического взаиморасположения исследованных восточнославянских групп. В результате этого можно заключить, что, несмотря на влияние естественного направленного отбора, система генов HLA сохраняет свое разнообразие, приуроченность различных вариантов её генов к определенным этническим группам и адекватно отражает геногеографию различных популяций, и, следовательно, может быть использована в популяционных исследованиях.
ВЫВОДЫ:
1. Самыми частыми аллельными вариантами DRB1 гена для всех изученных популяций являются: DRB1*07, *11, *13, *15, что соответствует западно-евразийскому типу распределения.
2. При попарном сравнении распределения специфичностей гена DRB1 между восточными украинцами и русскими выявлены достоверные различия в частотном содержании DRB1*11 и DRB1*16.
3. При попарном сравнении распределения аллелей гена DRB1 выявлено, что западные украинцы достоверно отличаются от русских частотным содержанием DRB1*11.
4. Сравнительный анализ представленных данных отмечает, что у всех популяций часто встречаемыми являются следующие трехлокусные гаплотипы: DRB1*11-DQA1*0501-DQB1*0301, DRB1*15-DQA1*0102-DQB1*0602-8, DRB1*01-DQA1*0101-DQB1*0501, DRB1*04-DQA1*0301-DQB1*0302 и DRB1*07-DQA1*0201-DQB1*0201
5. Отличительной особенностью является отсутствие или низкие частоты гаплотипов: DRB1*09-DQA1*0301-DQB1*0303 и DRB1*12-DQA1*0501-DQB1*0301
6. При попарном сравнении частот распределения трехлокусных гаплотипов (DRB1-DQA1-DQB1) восточных украинцев с русскими было установлено, что достоверными являются различия в частотном содержании DRB1*11-DQA1*0501-DQB1*0301 и DRB1*16-DQA1*0102-DQB1*0502/4.
7. Результаты многомерного статистического анализа выявили, что изученные популяции образуют два кластера»: «южный», включающий украинцев (Хмельницкая и Львовская области), и «северный», включающий белорусов (Витебская и Брестская области) и русских из Вологодской области.
8. При проведении многомерного кластерного анализа изученных популяций и некоторых народов Европы выявлено, что белорусы из Витебской и Брестской областей и украинцы из Хмельницкой и Львовской областей генетически близки с «центрально-европейскими» популяциями. Русские из Вологодской области сходны по HLA–профилю с «североевропейскими» и «центрально-европейскими» популяциями.
9. На примере восточных славян показано, что, несмотря на влияние естественного направленного отбора, система генов HLA адекватно отражает геногеографию этих этнических групп, и может быть использована в популяционных исследованиях.
Analysis of Y chromosome STR haplotypes in the European part of Russia reveals high diversities but non-significant genetic distances between populations Были исследованы Y-хромосомы 545 русских мужчин из Смоленской, Брянской, Ивановской, Липецкой, Пензенской, Рязанской, Орловской, Тверской, Вологодской, Тамбовской, Архангельской и Новгородской областей, т.е. по сути дела всего ареала образования великороссов.  Наиболее важные выводы: Whereas gradual change in the Y chromosome pool exists between Russia and the Slavic-speaking populations to the West, remarkable discontinuities were observed with neighbouring populations in the East, North and South. В то время как переход в рамках пула Y-хромосом между Россией и славяноязычными популяциями к западу от нее является постепенным, в отношении соседних популяций на востоке, севере и юге наблюдается примечательная обрывистость. No genetic substructure was found among the Russian populations. All pairwise comparisons were non-significant (p > 0.05). In contrast, significant variation between populations was observed in the comparison of Western Russia, treated as one homogeneous metapopulation, with neighbouring groups. Среди русских популяций не было обнаружено никакой генетической субструктуры. Все парные сравнения оказались незначительными (p > 0,05). В противоположность этому, существенная разница между популяциями была обнаружена при сравнении Западной России, рассматриваемой как одна однородная метапопуляция, с соседними группами. All pairwise Φst comparisons between Russia and these neighbours [Ukraine, Belarus, Lithuania, Latvia, Estonia, Finland, Siberia and the Caucasus region] (with the exception of Russia vs. Belarus) were significant with values ranging between 0.0089 (Russia vs. Poland) and 0.3688 (Russia vs. eight Altaic- and Uralic-speaking groups from Siberia). MDS plot based on pairwise Φst values shows a closely related core group consisting of Slavic-speaking populations (Russia, Ukraine, Belarus and Poland) with an elevated distance to Baltic populations and a large span to linguistically different groups from Estonia, Finland, Siberia and the Caucasus. Все парные сравнения по Φst между Россией и этими соседями [Украиной, Белоруссией, Литвой, Латвией, Эстонией, Финляндией, Сибирью и Кавказом] (за исключением России относительно Белоруссии) оказались существенными при разбросе значений от 0,0089 (Россия относительно Польши) до 0,3688 (Россия относительно восьми алтае- и уралоязычных групп из Сибири). Плот MDS на основании парных значений Φst представляет тесно связанную центральную группу, состоящую из славяноязычных популяций (России, Украины, Белоруссии и Польши), которая находится на более значительном расстоянии от балтийских популяций и на большом расстоянии от иноязычных групп из Эстонии, Финляндии, Сибири и Кавказа.
Наиболее важные выводы: Whereas gradual change in the Y chromosome pool exists between Russia and the Slavic-speaking populations to the West, remarkable discontinuities were observed with neighbouring populations in the East, North and South. В то время как переход в рамках пула Y-хромосом между Россией и славяноязычными популяциями к западу от нее является постепенным, в отношении соседних популяций на востоке, севере и юге наблюдается примечательная обрывистость. No genetic substructure was found among the Russian populations. All pairwise comparisons were non-significant (p > 0.05). In contrast, significant variation between populations was observed in the comparison of Western Russia, treated as one homogeneous metapopulation, with neighbouring groups. Среди русских популяций не было обнаружено никакой генетической субструктуры. Все парные сравнения оказались незначительными (p > 0,05). В противоположность этому, существенная разница между популяциями была обнаружена при сравнении Западной России, рассматриваемой как одна однородная метапопуляция, с соседними группами. All pairwise Φst comparisons between Russia and these neighbours [Ukraine, Belarus, Lithuania, Latvia, Estonia, Finland, Siberia and the Caucasus region] (with the exception of Russia vs. Belarus) were significant with values ranging between 0.0089 (Russia vs. Poland) and 0.3688 (Russia vs. eight Altaic- and Uralic-speaking groups from Siberia). MDS plot based on pairwise Φst values shows a closely related core group consisting of Slavic-speaking populations (Russia, Ukraine, Belarus and Poland) with an elevated distance to Baltic populations and a large span to linguistically different groups from Estonia, Finland, Siberia and the Caucasus. Все парные сравнения по Φst между Россией и этими соседями [Украиной, Белоруссией, Литвой, Латвией, Эстонией, Финляндией, Сибирью и Кавказом] (за исключением России относительно Белоруссии) оказались существенными при разбросе значений от 0,0089 (Россия относительно Польши) до 0,3688 (Россия относительно восьми алтае- и уралоязычных групп из Сибири). Плот MDS на основании парных значений Φst представляет тесно связанную центральную группу, состоящую из славяноязычных популяций (России, Украины, Белоруссии и Польши), которая находится на более значительном расстоянии от балтийских популяций и на большом расстоянии от иноязычных групп из Эстонии, Финляндии, Сибири и Кавказа. 

{kind=link}
Т.е. лишний раз доказано, что русские являются внутренне однородным славянским народом.
© 2011 г. В. В. Жиркова, С. А. Федорова, В. Л. Ахметова, Л. А. Животовский, Э. К. Хуснутдинова
Якутский научный центр комплексных медицинских проблем Российской академии медицинских наук,
Институт биохимии и генетики Уфимского научного центра Российской академии наук
Институт общей генетики им. Н.И. Вавилова Российской академии наук, Москва,
Проведен сравнительный анализ распределения частот аллелей шести STR-локусов (D3S1358, D16S539, THOI, D8S1179, LPL, HUMvWFII),применяемых в судебно-медицинской практике, в популяциях Якутии (три этногеографические группы якутов, эвенки, юкагиры, долганы, русские). По распределению частот пяти маркеров популяции русских достоверно отличаются от всех остальных популяций. Приведены значения совокупного дискриминирующего потенциала (PD) изученной системы локусов для каждой из изучаемых популяций. Уровень межпопуляционных генетических различий (FST) равен 0.005. На основе аллельного полиморфизма STR-локусов оценены филогенетические взаимоотношения между изученными популяциями и популяциями других регионов — Восточной Европы, Южной Сибири, Чукотки и Камчатки.
Ключевые слова: аутосомные микросателлитные локусы, популяции Якутии, генетическое разнообразие.
ALLELIC POLYMORPHISM OF SIX MICROSATELLITE DNA LOCI IN POPULATIONS OF SAKHA (YAKUTIA), by V. V. Zhirkova, S. A. Fedorova, V. L. Akhmetova, L. A. Zhivotovsky, E. K. Khusnutdinova, Yakut Research Center of Complex Medical Problems, Russian Academy of Medical Sciences, Yakutsk, 677010 Russia
Institute of Biochemistry and Genetics, Ufa Research Center, Russian Academy of Sciences, Ufa, 450054 Russia avilov Institute of Genetics, Russian Academy of Sciences, Moscow, 119991 Russia). The distribution of allele frequencies of six STR-loci (D3S1358, D16S539, THOI, D8S1179, LPL, HUMvWFII),used in forensic practice, was analyzed in populations of Sakha (Yakutia) (three ethnogeographical groups of Sakha (Yakuts), Evenks, Yukagirs, Dolgans, Russians). Significant differences were revealed between the Russians and all other populations by five markers. Values of total discriminative potential (PD) of studied system for each population were estimated. The level of interpopulation genetic differences (FST) was 0.005. On the basis of allelic polymorphism of STR-loci we estimated phylogenetic relationships between populations under study and populations of different regions — East Europe, South Siberia, Chukotka and Kamchatka.
Keywords:autosomal microsatellite loci, populations of Sakha (Yakutia), genetic diversity.
Исследование ДНК cчитается одним из наиболее актуальных методов анализа биологического материала в судебно-медицинских и криминалистических целях. Среди ДНК-маркеров различного типа, используемых в судебно-генетических исследованиях, предпочтение отдают аутосомным микросателлитам. Одну из основных проблем молекулярно-генетической идентификации личности и установления биологического родства представляет интерпретация полученных результатов, особенно в случае спорного отцовства/материнства, когда при исключении отцовства результаты не вызываю сомнений. Рассчитываемые параметры (индекс отцовства, вероятность отцовства и другие) основываются на популяционных частотах встречаемости выявленных у анализируемых индивидов аллелей.
На сегодняшний день общепризнано, что этнические группы различаются по частотам аллелей и генотипов отдельных ДНК-локусов. Это особенно актуально для географических областей, населенных разными этническими группами, включающими различные антропологические типы. При неправильном выборе референсной популяции величины вероятностей идентификации или биологического родства определяются с погрешностями и могут оспариваться при проведении судебной экспертизы как недостоверные. Результат анализа ДНК может не иметь юридической силы в том случае, если предварительно не установлена этническая принадлежность испытуемого, не проведены исследования в этнических группах и не учтены популяционные различия в частотах аллелей. Необходимо подчеркнуть, что анализ микросателлитных локусов до сих пор проводили главным образом в европейских популяциях. К настоящему времени изучено значительное число русских популяций из различных регионов Российской Федерации. В последние годы получен STR-характеристики популяций Волго-Уральского региона и Южной Сибири .
Цель настоящей работы — изучение полиморфизма шести аутосомных микросателлитных локусо (D3S1358, D16S539, THOI, D8S1179, LPL, HUM-vWFII)для создания референсной базы популяционно-генетических данных в семи популяциях коренного населения Якутии, необходимой при вычислении вероятностной статистики.
В работе использованы 317 образцов ДНК коренных жителей Республики Саха (Якутия). Коллекция составлена из материала Банка ДНК Отдела молекулярной генетики ЯНЦ КМП РАМН, собранного в экспедиционных выездах 2001—2006 гг. В выборку вошли неродственные индивиды, этническую принадлежность которых учитывали до третьего поколения. Всего проанализированы семь популяций: центральные якуты (n = 53), населяющие Лено-Амгинское междуречье (Чурапчинский, Усть-Алданский, Хангаласский, Амгинский, Мегино- Кангаласский, Намский улусы), вилюйские якуты (n = 49), проживающие в бассейне реки Вилюй (Сунтарский, Нюрбинский, Вилюйский, Верхне - вилюйский улусы), северные якуты (n = 53) (Верхоянский, Момский, Абыйский, Среднеколымский улусы), эвенки Жиганского, Усть-Майского, Оленекского улусов (n= 49), юкагиры Верхнеколымского и Нижнеколымского улусов ( n= 53), долганы Ана- барского улуса (n = 13), русскиe (n = 47) — жители населенных пунктов Мирнинского, Ленского, Алданского, Нерюнгринского улусов.
Суммарную ДНК выделяли из лимфоцитов периферической крови методом фенол-хлороформной экстракции с использованием протеиназы К. Матричные препараты ДНК амплифицировали методом ПЦР с помощью реагентов ТАПОТИЛИ (Государственный научный центр Российской Федерации “ГосНИИгенетика”, Москва). Амплификацию проводили на приборе Терцик (“ДНК-технология”, Россия). Продукты амплификации разделяли с помощью электрофореза в 10%-ном полиакриламидном геле с последующим окрашиванием бромистым этидием и серебром и визуализировали в УФ- свете системы гель-видеодокументации (“Vilber Lourmat”, Франция).
Статистический анализ данных (частоты аллелей и их ошибки, фактическая и теоретическая гетерозиготность, соответствие распределения генотипов равновесию Харди—Вайнберга) проводили с помощью пакета компьютерных програм Genepop (versio 3.3, 2001). Ожидаемую гетерозиготность и параметры информативности локусов: вероятность случайного совпадения генотипов двух неродственных индивидо (MF),вероятность дискриминации генотипов двух неродственных индивидо (PD) информативность полиморфизма данного локус (PIC) рассчитывали с помощью компьютерной программ PowerTyperExcel spreadsheet (“Promega” . Достоверность различий в распределении частот аллельных вариантов между популяциями оценивали с помощью точного теста программ RxC Факторный анализ с использованием метода главных компонент проводили с помощью программ POPSTR любезно предоставленно H. Harpending.
РЕЗУЛЬТАТЫ И ОБСУЖДЕНИЕ
В табл. 1 представлены данные о распределении частот аллелей шести STR-локусов (D3S1358, D16S539, LPL, THOI, D8S1179, HUMvWFII) в семи популяциях коренного населения Якутии.
Микросателлит D3S1358 расположен на хромосоме 3p21.3. В разных популяциях найдены 12 различных аллелей этого маркера размером от 99 до 147 п.н., включающих от 8 до 20 повторов ТСТА. У русских и у северных якутов выявлено шесть аллелей маркера D3S1358, у долган встречаются только четыре аллеля, во всех остальных популяциях обнаружено по пять аллелей. Из них наиболее распространен аллель с числом повторов 15, однако у долган и русских преобладает аллель с 16 повторами. По-видимому, высокая частота аллеля 16 в популяции долган свидетельствует о значительном содержании европеоидного компонента в их генофонде — мужской генофонд долган отличается высокой частотой западноевразийских линий (26%) .
В выборке вилюйских якутов аллель 14 не найден. Самый высокомолекулярный аллель 19 обнаружен у северных и вилюйских якутов, а также у русских. В целом эти данные согласуются с результатами, полученными для народов Южной Сибири, Волго-Уральского региона, где также наблюдается одномодальное распределение частот аллелей с преобладанием аллеля 15. Индексы фактической и теоретической гетерозиготности представлены в табл. 2. Наиболее высокие значения наблюдаемой гетерозиготности (Hobs= 0.750 и 0.702) выявлены в популяциях долган и русских. Показатель теоретической гетерозиготности в среднем для семи популяций коренного населения Якутии составил Hexp= 0.660.
Микросателлит D16S539 расположен на хромосоме 16 в районе q24. Этот маркер характеризуется наибольшим внутрипопуляционным разбросом аллелей и считается одним из наиболее вариабельных. В различных популяциях найдены аллели с числом повторов от пяти (94 п.н.) до 16 (138 п.н.), в том числе “промежуточные” аллели 9.3, 11.3, 12.1, 12.2, 13.3, 14.3 . Во всех популяциях отмечено бимодальное распределение с первым пиком аллеля 9, основного во всех изученных популяциях, и вторым пиком в области аллелей 11—13. Такая картина характерна для народов Волго-Уральского региона, Южной Сибири, Центральной России, Северной Кореи, Чукотки и Камчатки . В наших популяциях мы обнаружили восемь аллелей. В выборках северных якутов, юкагиров и долган найдено по шесть аллелей. Аллели 9, 11, 12, 13 обладают наибольшими частотами встречаемости во всех популяциях. Аллель 8 не найден у юкагиров и долган. Уровень фактической гетерозиготности (Hobs) варьирует от 0.68 у эвенков до 0.846 у долган. В целом в популяциях коренного населения Якутии среднее значение наблюдаемой гетерозиготности по локус D16S539 (Hobs 0.776) несколько выше ожидаемой (Hexp 0.760).
Микросателлит LPL расположен в интроне 6 гена липопротеидлипазы (хромосомная локализация 8p22). Этот маркер включен в состав коммерческих наборов “Promega Corp.”. В различных популяциях представлены аллели этого маркера с числом повторов от семи (105 п.н.) до 16 (141 п.н.) . Нами выявлены восемь различных аллелей маркера LPL (от трех до шести в разных группах). Распределение частот аллелей во всех популяциях в целом характеризуется одномодальностью с преобладанием частоты аллеля 10. При этом у центральных и у вилюйских якутов обнаружен аллель 7, которого нет в выборках северных якутов, юкагиров, долган и русских. Самый высокомолекулярный аллель 14 встречается только в выборке вилюйских якутов. В силу высокой частоты аллеля 10 (как правило, более 40%) LPL выделяется наименьшим уровнем гетерозиготности среди изучаемых локусов. Для всех популяций коренного населения Якутии, кроме русских, характерны выраженный дефицит гетерозигот и невысокий уровень фактической гетерозиготности:
0.46 < Hobs lt; 0.56.
Микросателлит THOI расположен в первом интроне гена тирозингидроксилазы, локализованного на коротком плече хромосомы 11. Описано восемь аллелей этого локуса с числом повторяющихся единиц от пяти до 11. В выборках юкагиров выявлено семь аллелей маркер THOI, у русских — шесть аллелей, у долган только четыре, а во всех остальных популяциях — пять аллелей. Из них в популяциях центральных, вилюйских якутов, эвенков и юкагиров наиболее распространен аллель с семью повторами. Согласно, аналогичное преобладание аллеля 7 характерно для популяций бурятов, сойотов, хакасов, чукчей, коряков и хамниганов. В популяциях русских, долган и северных якутов с наибольшей частотой встречается аллель 9.3, характерный также для народов Волго-Уральского региона, Центральной России, Северной Кореи . Индексы наблюдаемой гетерозиготности (Hobs варьировали от 0.740 в популяции северных якутов до 0.880 у эвенков. Среднее значение фактической гетерозиготности в изученных группа (Hobs=0.810) оказалось выше средней теоретической гетерозиготности (Hexp= 0.766).
Микросателлит D8S179 расположен на хромосоме 8q24.1—q24.2. В пяти популяциях обнаружили восемь различных аллелей этого маркера. В выборках юкагиров и долган найдено шесть и пять аллелей соответственно. Во всех популяциях отмечен унимодальное распределение аллелей с максимальной частотой аллеля 13. В целом, наши данные согласуются с результатами, полученными для народов Южной Сибири, Волго-Уральского региона, Центральной России, Северной Кореи, Чукотки и Камчатки, где также наблюдается одномодальное распределение частот аллелей с преобладанием аллеля 13 . Показатель теоретической гетерозиготности в среднем для семи популяций коренного населения Якутии составил Hexp= 0.650.
Микросателлит HUMvWFII находится в 3'-концевой части полиморфного участка интрона 40 гена фактора Виллебранда. Этот маркер входит в состав идентификационных тест-систем в России. В разных популяциях найдены девять различных аллелей с восемью (150 п.н.)—16 (182 п.н.) повторами, причем у европеоидов аллель 8 достаточно редкий. Этот полиморфизм изучен в популяциях якутов, эвенков, юкагиров и долган. В выборках центральных, северных якутов, юкагиров обнаружено семь различных аллелей микросателлит HUMvWFII, у эвенков и вилюйских якутов по шесть аллелей, у долган — четыре. Во всех популяциях наблюдается унимодальное распределение аллелей с максимальной частотой аллеля 11, кроме выборки долган, где наибольшая частота отмечена для аллеля 12. При этом во всех популяциях наибольшие частоты имеют аллели 11, 12 и 13. У вилюйских якутов, эвенков и долган отсутствует аллель 10. Наиболее высокие значения наблюдаемой гетерозиготност (Hobs= 0.780 и 0.760) выявлены в популяциях эвенков и центральных якутов. Показатель теоретической гетерозиготности в среднем в шести популяциях коренного населения Якутии состави Hexp= 0.690.
Всего в изученных нами популяциях выявлено 45 аллелей. В отдельных выборках число аллелей варьирует от 26 (у долган) до 40 (у русских). Анализ шести судебных локусов показал высокий уровень разнообразия среди популяций Якутии. В изученных выборках обнаружены по семь—восемь аллелей четырех из шести маркеров (восемь D16S539, D8S1179 и семь HUMvWFII и THOI).
Во всех популяциях не найдено существенных отклонений от равновесия Харди—Вайнберга.При попарном сравнении частот аллелей маркеров D3S1358, LPL, THOI, D8S1179 HUMvWFI становлены достоверные различия между русскими и остальными популяциями. Анализ гетерогенности микросателлитных маркеров D16S539, HUMvWFI выявил достоверные отличия долган от всех остальных популяций. Показано, что по распределению частот аллелей микросателлит D8S1179 популяция юкагиров достоверно отличается от всех остальных популяций.
Межпопуляционные различия
Для оценки генетических различий между популяциями, а также между элементарными популяциями внутри одной крупной популяции широко используется показатель Райт (FST) Средняя субэтническая дифференциация популяций Якутии составил FST =0.016 (табл. 2). Только по маркер D16S539 популяции имеют очень малые различи (FST= 0.002). Наибольший вклад в генетическую дифференциацию популяций Якутии внес марке D8S1179 FST =0.030). Кроме того, степень различий между коренными этносами Якутии по шести ДНК-маркерам ядерного геном (FST= 0.005) сравнима со степенью генетической дифференциации по частотам гаплогрупп мтДН (FST= 0.009) и намного ниже, чем тот же показатель по частотам гаплогруп Y-хромосомы — 0.207. Ранее было установлено, что различия в степени дифференцированности мужского и женского генофондов населения Якутии обусловлены эффектом основателя в популяциях якутов и эффектом патрилокальности. По - видимому, системы аутосомных микросателлитных маркеров и мтДНК в целом дают более объективное представление о степени генетических различий между популяциями Якутии, чем Y-хромосома.Степень различий между генофондами этногео графических групп якутов — центральных, вилюйских, северных — оказалась (FST= 0.002) ниже, чем уровень генной дифференциации у удмуртов (FST= 0.004), рассчитанный по 10 локусам ядерного генома, и у башкир (FST= 0.006), рассчитанный по четырем локусам ядерного генома. Этот показатель также более сравним со степень генетической дифференциации по гаплогруппам мтДН (FST= 0.008), чем по Y-хромосоме (FST= 0.044).
Генетические взаимоотношения между популяциями
Генетические взаимоотношения между популяциями Якутии и других регионов Российской Федерации оценивали с использованием метода главных компонент (РС-анализ), основанном на частотах аллелей микросателлитных локусов в популяциях. На рис. 1 показано положение популяций Якутии в пространстве двух главных компонент, описывающих 62% вариабельности частот аллелей. Популяции якутов и эвенков образуют отдельный компактный кластер вследствие высоких частот аллеля 10 локуса LPL, аллеля 13 локуса D8S1179, аллеля 15 локуса D3S1358. Популяция юкагиров дистанцирована от них из-за повышенной частоты аллеля 10 локуса THOI, аллеля 15 локуса D8S1179. Популяция русских, генофонд которых характеризуется высокими частотами аллелей 11, 12 локуса LPL, аллеля 12 локуса D8S1179, аллеля 6 локуса THOI, аллеля 18 локуса D3S1358, расположена отдельно. В целом, характер кластеризации популяций в пространстве двух главных компонент по частотам аллелей микросателлитных локусов сходен с кластеризацией по частотам гаплогрупп мтДНК и Y-хромосомы.четко разделяются по первой компоненте на два кластера — первый образован европеоидными популяциями и тофаларами, второй — монголоидными популяциями Азии. В этом случае аллели, имеющие более высокую частоту в европеоидных популяциях, — это аллель 6 локуса THOI, аллель 18 локуса D3S1358, аллель 14 локуса D8S1179. Монголоидные популяции характеризуются высокими частотами аллеля 9 локуса D16S539, аллеля 15 локуса D3S1358, аллеля 7 локуса THOI, аллеля 13 локуса D8S1179. По вариабельности частот микросателлитных локусов популяции Якутии группируются вместе с популяциями Южной Сибири, отдалены от популяций Чукотки и Камчатки, что соответствует данным анализа мтДНК.
Изученные нами аутосомные микросателлитные локусы могут быть использованы в криминалистической и судебной практике Республики Саха (Якутия) и других регионов России, а именно, при молекулярно-генетическом определении личности, а также при установлении биологического родства.
К ПРОБЛЕМЕ ПРОИСХОЖДЕНИЯ МОНГОЛОИДНОГО КОМПОНЕНТА МИТОХОНДРИАЛЬНОГО ГЕНОФОНДА СЛАВЯН
© 2008 г. Б. А. Малярчук, М. А. Перкова, М. В. Деренко
Институт биологических проблем Севера Дальневосточного отделения Российской академии наук,
Представлены данные о рестрикционном полиморфизме митохондриальной ДНК (мтДНК) в чешской популяции (n = 279). Установлено, что в структурном отношении митохондриальные генофонды чехов и других славянских народов (русских, поляков, словенцев, боснийцев) практически не различаются. Частота восточноевразийских (монголоидных) линий мтДНК у чехов составила 1.8%. Проводится анализ распространенности восточноевразийских линий мтДНК в популяциях Европы, относящихся к различным этнолингвистическим группам. Выявлены различия по частоте подобных линий мтДНК в различных группах славян: от 1.2 и 1.6% соответственно у южных и западных славян до 1.3-5.2% у восточных славян - русского населения Восточной Европы. Наиболее высокая частота монголоидного компонента зарегистрирована в митохондриальных генофондах русского населения Русского Севера и Северо-Западного региона, что может объясняться ассимиляцией славянами североевропейских финно-угорских народов в процессе формирования русского населения этих регионов. Обсуждается проблема происхождения монголоидного компонента в генофондах различных групп славян.
Генетические аспекты истории формирования населения Европы исследованы еще недостаточно, хотя различным проблемам в этой области уже посвящен ряд исследований . Среди них наиболее активно развиваются исследования полиморфизма высокополиморфных генетических систем - митохондриальной ДНК (мтДНК) и Y-хромосомы, наследуемых по одной из родительских линий. С помощью анализа полиморфизма мтДНК изучены популяции восточных, западных и южных славян . Анализ данных об изменчивости мтДНК в европейских популяциях, включая славянские, показал, что славяне характеризуются единством происхождения, центральное положение среди них занимают западные славяне, а степень генетических различий между группами славян определяется в значительной мере степенью смешения с дославянским населением современного этнического ареала славян, а также интенсивностью их взаимодействия с соседними народами . Последний вывод следует из того факта, что соседи русских - западнофинские популяции - характеризуются относительно высоким генетическим сходством с русскими, германские популяции - с западными славянами, а балканские - с южными славянами .
Между тем не все группы славян охарактеризованы в одинаковой мере. Довольно детально исследовано разнообразие митохондриальных генофондов южных и восточных славян, а западные славяне представлены практически одной этнической группой - поляками . Чехи и словаки, также относящиеся к числу западнославянских народов, изучены явно недостаточно . Целью настоящей работы является изучение разнообразия мтДНК в чешской популяции и сравнительный анализ распределения групп мтДНК у славян и соседних по отношению к ним народов Европы.
МАТЕРИАЛЫ И МЕТОДЫ
Сбор биологического материала (цельная кровь) осуществлялся на базе терапевтических отделений больниц от индивидуумов, не отягощенных наследственной патологией. Исследуемая выборка (n = 279) представлена индивидуумами чешской национальности - уроженцами различных районов Чешской Республики.
Материалом для исследований служила геномная ДНК, выделенная из крови с использованием стандартных методов, включающих лизис клеток протеиназой K (“Sigma”, USA) в присутствии 1% додецилсульфата натрия, очистку ДНК фенолом/хлороформом и осаждение ДНК этиловым спиртом.
Скрининг полиморфных сайтов, определяющих основные группы типов мтДНК, распространенных в популяциях Евразии (табл. 1), проводился посредством анализа участков мтДНК, амплифицированных в полимеразной цепной реакции, с использованием праймеров, предложенных в работах . Рестрикционные фрагменты фракционировались электрофоретически в 8%-ных полиакриламидных гелях. Для детекции ДНК использовалась окраска гелей бромистым этидием с последующей визуализацией ДНК в УФ-свете. Полиморфизм учитывался по наличию (+) и отсутствию (-) сайтов рестрикции.
Идентификацию типов мтДНК проводили на основании существующей классификации мтДНК в популяциях человека . В соответствии с классификацией группы мтДНК, за исключением группы HV, обозначаются латинским однобуквенным кодом. Учитывая рекомендации, изложенные в недавней работе , кластер pre-HV обозначается как R0, (pre-HV)1 - как R0a, pre-V - как HV0, pre-V1 - как HV0b и pre-V2 - как HV0a.
Для идентификации образцов мтДНК, не классифицированных с помощью схемы анализа, показанной в табл. 1, нами проводился скрининг дополнительных маркеров, определяющих группы L1 и L2 (в пределах кластера L) и N9a (в пределах кластера N, за исключением R). Для идентификации образцов в пределах групп L1 и L2 использовали схему рестрикционного анализа, представленную в работе . Для определения группы N9a анализировали JasI-полиморфизм участка 5416-5419. Типы мтДНК, характеризующиеся вариантом +5416JasI, определяли как N9a-типы .
Достоверность межпопуляционных различий по частотам групп типов мтДНК оценивали с помощью точного теста на популяционную дифференциацию . Индексы разнообразия мтДНК в популяциях и значения F-статистик рассчитывали с помощью компьютерных программ пакета Arlequin 2.0 , предназначенного для анализа молекулярной изменчивости и генетической структуры популяций.
РЕЗУЛЬТАТЫ И ОБСУЖДЕНИЕ
Анализ изменчивости мтДНК у чехов показал, что их генофонд характеризуется типично европейской композицией групп и подгрупп мтДНК. Наиболее представленными у них, как и в других славянских популяциях, являются кластеры H, U, T, J (табл. 2). Подавляющее большинство кластеров мтДНК, выявленных у чехов, являются западноевразийскими по происхождению. Частота восточноевразийских (монголоидных) линий мтДНК у них составила 1.8% (группы A, N9a, M). Обнаружена также африканская линия (частота 0.4%), относящаяся к группе L2a, маркированной вариантом +13803HaeIII. В структурном отношении митохондриальные генофонды исследованных групп славян очень сходны по композиции групп и подгрупп мтДНК (табл. 2). Анализ генетической дифференциации популяций показал отсутствие межпопуляционных различий во всех парах сравнений как с помощью F-статистики (Fst = 0, р = 0.68), так и с использованием точного теста (р = 0.3 ± 0.08).
Анализ изменчивости мтДНК в популяциях Европы показал, что в генофондах различных славянских популяций практически всегда присутствуют линии ДНК, имеющие восточноевразийское происхождение (табл. 3). Однако неясно, чем это обусловлено: входил ли достаточно разнообразный монголоидный компонент в состав предкового генофонда славян или же его появление в генофондах различных групп славян имеет “накопительный” характер, т.е. связано с постепенной ассимиляцией монголоидных линий ДНК в результате взаимодействия между населением Восточной Европы и Азии в различные эпохи?
Согласно лингвистическим, археологическим и палеогеографическим данным, распад балто- славянской языковой общности произошел примерно во второй половине III тыс. до н.э., что привело к появлению протобалтов и протославян . Анализ изменчивости мтДНК у славян и балтов показал, что их генофонды различаются по частоте восточноевразийского компонента (табл. 3). Низкая частота монголоидных вариантов мтДНК у латышей и литовцев свидетельствует о том, монголоидный компонент, по всей видимости, не был характерен для балто-славянского протогенофонда. Таким образом, правомочно предположение о том, что процесс накопления монголоидных линий мтДНК у славян и их предков интенсифицировался лишь на протяжении последних четырех тысячелетий.
Исследования изменчивости мтДНК у русского населения Восточной Европы показали, что региональные группы русских популяций различаются по частоте и композиции групп мтДНК, имеющих восточноевразийское происхождение. Данные табл. 3 показывают, что частота монголоидного компонента повышается в северном направлении. Наиболее высокие частоты монголоидного компонента характерны для русского населения Русского Поморья и Северо-Западного региона, однако популяции этих регионов различаются по композиции групп мтДНК.
Известно, что в процессе формирования русского населения большое значение имела ассимиляция коренного дославянского населения Восточной Европы собственно славянами. Поэтому межрегиональные различия между группами русских могут быть обусловлены генетическими особенностями ассимилированных популяций. Установлено, что для североевропейских финно-угорских популяций (таких, как саамы, финны, карелы) характерно присутствие главным образом двух групп мтДНК - Z и D5 . Исходя из результатов филогеографического анализа предполагается, что эти группы мтДНК могли быть привнесены на север Европы из Азии еще в эпоху раннего неолита . Поэтому высокая частота групп Z и D5 у русских поморов, и в меньшей степени у русского населения Северо-Западного региона, обусловлена, по-видимому, ассимиляцией главным образом североевропейских финно-угорских народов. Между тем спектр восточноевразийских групп мтДНК у населения большинства регионов европейской части России, включая ее северо-западную часть, значительно шире, чем у населения Русского Поморья . Это свидетельствует о том, что ассимиляции был подвержен круг других популяций, генетически более сходных с такими современными восточноевропейскими народами, как мордва, марийцы, удмурты и коми. Усложнение структуры монголоидного компонента генофондов этих народов, по всей видимости, обусловлено долговременным взаимодействием с народами Сибири и Центральной Азии. Одним из главных периодов усиления этого воздействия является раннее средневековье, когда волны миграций целого ряда степных народов (гунны, авары, булгары, монголы) прокатились по Восточной Европе. Так, в VI веке н.э. авары достигли даже территорий Центральной Европы (Валахии, Паннонии, Трансильвании и Богемии), где ими был образован Аварский каганат, существовавший влоть до IX в. н.э. Предполагается, что после распада Аварского каганата народы, входившие в его состав, были ассимилированы племенами славян . Таким образом, присутствие ощутимой частоты восточноевразийских линий мтДНК в генофондах некоторых популяций западных и южных славян (особенно на территории бывшего Аварского каганата), по всей видимости, является следствием этого процесса.
Что касается населения Восточной Европы, то антропологические данные показывают, что лесная полоса Восточной Европы была зоной интенсивного смешения народов . Предполагается, что формирование монголоидного комплекса признаков происходило не позднее верхнего палеолита, в связи с чем восточносибирские популяции могли располагать значительным запасом времени для миграций в Восточную Европу . Невыясненным остается вопрос о количестве этих миграций, так как на северо-западе Восточной Европы монголоидный компонент наблюдается 10-8 тыс. лет назад, в составе днепро-донецких племен 7-6 тыс. лет назад, на территории Ивановской области (Сахтыш) 6-5 тыс. лет назад . Результаты анализа изменчивости мтДНК в русских популяциях хорошо согласуются с антропологическими данными, поскольку указывают на существенные различия по частоте монголоидных линий мтДНК между русским населением Русского Севера, Северо-Запада и центральных/южных районов европейской части России (табл. 3). Надеемся, что в перспективе станут возможными исследования хронологии формирования разнообразия монголоидного компонента, присутствующего в генофондах русского населения и в других популяций славян.
Следы балтийских славян В генофонде русского населения Восточной Европы.
Борис
Аркадьевич Малярчук, доктор биологических наук,
заведующий лабораторией генетики Института биологических проблем Севера Дальневосточного отделения РАН (Магадан). Область научных интересов - использование методов молекулярной генетики в
решении проблем происхождения народов, этногенез славян,
мутагенез ДНК.
Абстракт
Анализ изменчивости
митохондриальной ДНК (мтДНК) и Y-хромосомы в популяциях русского
населения европейской части России показал, что русские характеризуются низким
уровнем генетической дифференциации как по материнским линиям
мтДНК, так и по отцовским линиям Y-хромосомы.
Более того, генетические различия между русскими
популяциями вообще исчезают, если их сгруппировать в соответствии с диалектным
членением русского языка или по данным антропологии. Между тем, анализ
изменчивости мтДНК в популяциях северной части Европы показал,
что некоторые русские популяции Северо-Западного региона России (Псков и
Великий Новгород) и поляки Северо-Восточной Польши (Сувалки) достоверно
отличаются от соседних популяций славянского (русские и поляки), балтского (литовцы) и угро-финского (эстонцы, карелы, финны) происхождения.
Полученные генетические данные позволяют рассматривать псковско-новгородское
русское население в качестве отдельной славянской группировки в составе современных восточных славян.
В последние годы значительно
усилился интерес к изучению генетических систем, наследуемых
по одной из родительских линий - митохондриальной ДНК (мтДНК), наследуемой по материнской линии, и Y-хромосомы,
наследуемой по отцовской линии. Совместный анализ этих
генетических систем позволяет изучать генетическую структуру популяций и
реконструировать историю формирования генофондов с учетом вклада мужских и женских линий ДНК. Результаты подобных исследований
русского населения европейской части России показали, что
межпопуляцион-ные различия между русскими популяциями не
очень велики (Malyarchuk et al.,
2004; Малярчук, Деренко, 2008; Корниенко и др., 2009). Необходимо отметить, что
добавление в анализ популяций, представляющих
2008)
. Однако поморы отличаются высоким
генетическим сходством с финно-угорским населением северной части Европы и высокой степенью подразделенности генофонда, вызванной влиянием дрейфа (Khrunin et al.,
2009) , и поэтому не могут рассматриваться в качестве
типично русского населения - такого,
например, как население центральных и южных районов европейской части России.
Нами проведен анализ изменчивости 11 микросателлитных участков
(локусов) Y-хромосомы (DYS19, DYS385a,b, DYS389I,
DYS389II, DYS390, DYS391, DYS392, DYS393, DYS437, DYS438, DYS439) в популяциях русского населения Калужской, Тульской, Владимирской,
Нижегородской, Белгородской, Орловской, Саратовской, Новгородской (г. Великий
Новгород и с. Волот), Псковской, Ростовской областей,
Ставропольского и Краснодарского краев (всего
проанализировано 594 человека). Сравнительный анализ
показал наличие низкого, но статистически значимого, уровня межпопуляционных генетических различий. Показатель генетической
дифференциации FST
оказался равен 0.52% при достоверности различий на уровне P =
0.03. Для выявления возможных генетических различий нами проведен
анализ популяций, сгруппированных в соответствии с диалектным членением
русского языка на территории европейской части России и на основании
данных антропологии. Диалектным объединениям русского языка
отводится большая роль в изучении внутриэтнической дифференциации русского
народа (Захарова, Орлова, 1970). Вместе с тем, данные антропологии также свидетельствуют о том, что в этнотерриториальном
делении русского населения до сих пор отражаются
антропологические характеристики летописных племен
славян (Алексеева, 1973), и поэтому антропологический критерий вполне может
быть использован для определения состава анализируемых
групп популяций современного русского населения. Таким образом, популяции объединяли в группы следующим образом
(табл. 1):
(А)
по говорам и наречиям русского языка: южное наречие (Белгород, Орел,
Калуга, Тула), северное наречие (Ярославль), среднерусские говоры (Великий Новгород, Волот, Псков, Владимир, Нижний
Новгород);
(Б) по диалектным зонам: юго-западная зона (Белгород, Орел, Калуга,
Тула), северозападная зона (Великий Новгород, Волот, Псков), северо-восточная зона
(Владимир, Нижний Новгород, Ярославль);
(В) в
соответствии с данными антропологии: предполагаемые потомки словен
новгородских и псковских кривичей (Великий Новгород,
Волот, Псков), кривичей (Владимир, Нижний Новгород, Ярославль), вятичей (Калуга, Тула), северян (Орел, Белгород).
Проведенный анализ показал, что, несмотря
на более высокое значение межгрупповых различий при группировании
популяций в соответствии с распределением говоров и наречий русского языка в
сравнении с картой диалектных зон, в обоих случаях
межпо-пуляционные различия недостоверны (табл. 1).
Это свидетельствует об отсутствии генетических различий по распределению
линий Y-хромосомы
в группах русских популяций, выделяемых на основании данных лингвистики. Такой же вывод следует и из результатов
анализа дифференциации русских популяций, сгруппированных с учетом
данных антропологии (табл. 1).
Нами проведен также анализ
изменчивости нуклеотидных последовательностей гипервариабельного сегмента 1 (ГВС1) мтДНК в русских популяциях Тульской,
Калужской, Владимирской, Ярославской, Псковской, Новгородской (г. Великий Новгород и с. Волот), Белгородской,
Нижегородской, Орловской, Саратовской, Курской и Костромской областей и Ставропольского края (всего проанализировано
913 человек. В). Значение межпо-пуляционной дифференциации FST составило 0.35% (P = 0.0007). Для дальнейшего анализа популяции объединяли в группы
следующим образом:
(A) по
говорам и наречиям русского языка: южное наречие (Белгород, Орел,
Калуга, Тула, Курск), северное наречие (Ярославль, Кострома), среднерусские
говоры (Великий Новгород, Волот, Псков, Владимир, Нижний Новгород);
(Б) по диалектным зонам: юго-западная зона (Белгород, Орел, Калуга,
Тула, Курск), северо-западная зона (Великий Новгород, Волот, Псков),
северо-восточная зона (Владимир, Нижний Новгород, Ярославль,
Кострома);
(B) в
соответствии с данными антропологии: предполагаемые потомки словен новгородских
и псковских кривичей (Великий Новгород, Волот, Псков), кривичей
(Владимир, Нижний Новгород, Ярославль, Кострома), вятичей (Калуга, Тула),
северян (Орел, Белгород, Курск). Результаты этого исследования также показали отсутствие
статистически значимой дифференциации между русскими популяциями,
сгруппированными в соответствии с данными лингвистики и
антропологии
(табл. 1).
Ранее отмечалось некоторое
своеобразие митохондриального генофонда русского нанаселения Северо-Западного региона России
(Лункина и др., 2004; Малярчук, Деренко, 2006; Grzybowski et al., 2007). В связи с этим нами
проведен анализ данных об изменчивости нуклеотидных
последовательностей ГВС1 мтДНК в популяциях Северо-Западного
региона России (Великий Новгород, Волот и Псков) в сравнении с другими
славянскими популяциями России (русские) и Польши
(поляки), а также популяциями соседних народов: балтских (литовцы) и
угро-финских (эстонцы, карелы и финны). Исследование показало,
что степень дифференциации проанализированных популяций низка - FST = 0.38%, P = 0. Однако некоторые популяции
Северо-Западного региона России (Великий Новгород и Псков) и Северо-Восточной
Польши (Сувалки) демонстрируют очень высокое генетическое сходство. В табл. 2 показаны результаты иерархического анализа изменчивости мтДНК в группах популяций. Видно, что при группировании популяций в соответствии с их этнолингвистической характеристикой (группа русских, группа поляков и группа балтского и угро-финского происхождения) межгрупповые различия очень низки и недостоверны. Однако выделение отдельной группы популяций, включающей северо-западных русских (Великий Новгород и Псков), северо-восточных поляков (Сувал-ки) и популяции балтского и угро-финского происхождения, приводит к достоверному увеличению степени межгрупповой дифференциации (0.15%). Еще большие различия наблюдаются, если отдельная группа представлена только северо-западными русскими и северо-восточными поляками. Это свидетельствует о своеобразии генетической структуры этих популяций по отношению как к остальным славянским популяциям (русским и полякам; 0.52%), так и к популяциям балтско-го и угро-финского происхождения (0.76%). Анализ генофондов северо-западных русских и северо-восточных поляков в сравнении с их непосредственными соседями - литовцами и эстонцами - также показывает достоверные различия между ними (0.73%). Таким образом, генетическое своеобразие русского населения Северо-Западного региона России и Северо-Восточной Польши не может быть объяснено исключительно лишь их сходством с балтским и угро-финским населением, а является следствием появления уникальной генетической композиции, сложившейся, по-всей видимости, в результате взаимодействия трех генофондов - славянского, балтского и угро-финского. Анализ структуры митохон-дриального генофонда популяций Великого Новгорода, Пскова и Сувалок показал наличие лишь одного генетического компонента -гаплогруппы U5a, которая распространена в этих популяциях с более высокой частотой (в среднем, 16%), чем в соседних славянских, балтских и угро-финских популяциях, где ее частота, в среднем, составляет 7%. Интересной особенностью генофондов популяций Великого Новгорода, Пскова и Сувалок является также присутствие (в среднем, с частотой 2.2%) гаплотипов митохондриальных групп R1 и R2, которые крайне редки в соседних популяциях.
Генетические параллели между населением Северо-Западной России и Северо-Восточной Польши
Обратим внимание на любопытное исследование генофондов ряда русских, польских, литовских и западно-финнских популяций: "Следы балтийских славян в генофонде русского населения Восточной Европы", Б.Малярчук, 2009.По результатам настоящего и предшествующих исследований центрально- и восточно-европейских генофондов автор формулирует следующие тезисы:
1. Русские характеризуются низким уровнем генетической дифференциации как по материнским линиям мтДНК, так и по отцовским линиям Y-хромосомы.2. Несмотря на более высокое значение межгрупповых различий при группировании популяций в соответствии с распределением говоров и наречий русского языка в сравнении с картой диалектных зон, в обоих случаях межпопуляционные различия недостоверны. Это свидетельствует об отсутствии генетических различий по распределению линий Y-хромосомы в группах русских популяций, выделяемых на основании данных лингвистики. Такой же вывод следует и из результатов анализа дифференциации русских популяций, сгруппированных с учетом данных антропологии.3. Анализ изменчивости мтДНК в популяциях северной части Европы показал, что некоторые русские популяции Северо-Западного региона России (Псков и Великий Новгород) и поляки Северо-Восточной Польши (Сувалки) достоверно отличаются от соседних популяций славянского (русские и поляки), балтского (литовцы) и угро-финского (эстонцы, карелы, финны) происхождения.4. Полученные генетические данные позволяют рассматривать псковско-новгородское русское население в качестве отдельной славянской группировки в составе современных восточных славян.5. Генетическое своеобразие русского населения Северо-Западного региона России и Северо-Восточной Польши не может быть объяснено исключительно лишь их сходством с балтским и угро-финским населением населением, а является следствием появления уникальной генетической композиции, сложившейся, повсей видимости, в результате взаимодействия трех генофондов — славянского, балтского и угро-финского.6. Анализ структуры митохондриального генофонда популяций Великого Новгорода, Пскова и Сувалок показал наличие лишь одного генетического компонента — гаплогруппы U5a, которая распространена в этих популяциях с более высокой частотой (в среднем, 16%), чем в соседних славянских, балтских и угро-финских популяциях, где ее частота, в среднем, составляет 7%. Интересной особенностью генофондов популяций Великого Новгорода, Пскова и Сувалок является также присутствие (в среднем, с частотой 2.2%) гаплотипов митохондриальных групп R1 и R2, которые крайне редки в соседних популяциях.
Географическое расположение сходных популяций отмечено на следующей карте: Сходные популяции: Сувалки (wiki), Великий Новгород, ПсковК процитированному должен добавить следующее. Предшествующие исследования генофондов восточно-европейских народов выявили значительное сходство между поляками, белорусами и русскими в целом — местами даже более значительное, чем между русскими и украинцами. Вместе с этим, по отцовским линиям Y-хромосомы белорусы обнаружили близость не только с поляками, но и с далёкими лужицкими сербами [Балановский, Тегако, 2008]. Такое положение вещей хорошо иллюстрируют карты расстояний на основании данных по Y-хромосоме: от русских популяций, от белорусов и украинцев [Балановский, Балановская, 2008].По результатам автор касается истории и лингвистики, где находит параллель в отмеченных рядом историков следах колонизации областей Северо-Западной России большой группой балтийских славян — выходцев из Южной Прибалтики и Польского Поморья — в 5-6 вв. н.э.При этом речь идёт именно о появлении большой массы славянского населения в Приильменье и верховьях Западной Двины, которое констатируется и археологами, и лингвистами, и антропологами. В то же время, распространённой точкой зрения на появление славян на юго-западном побережье Балтики — предположительно, исходном ареале миграции — является мнение о вытеснении оттуда германцев около 4-5 вв. н.э. и появлении достоверно славянской суковско-дзедзицкой (wiki) археологической культуры. По всей вероятности, вытеснившая германцев группа славян была настолько многочисленна, что буквально за несколько столетий заселила не только свободившиеся земли, но и — морским путём — северо-западные области будущей Руси. Без теории пассионарности за авторством Л.Н.Гумилёва здесь обойтись не удастся.Если абстрагироваться от поиска исторических параллелей и вернуться к популяционной генетике, то указанный сценарий является далеко не единственным. Нам достоверно известен только факт сходства генофондов поляков, русских и белорусов, а также существование внутри них примечательной группы популяций Сувалки-Псков-Новгород, чего самого по себе недостаточно для заключения об имевшей место миграции с запада на восток или с востока на запад. Не менее вероятным выглядит «автохтонный» сценарий, в соответствии с которым указанная группа популяций некогда находилась в пределах единого ареала и была разделена лишь позже. За отсутствием откровенно противоречивых фактов возможность подобного процесса отвергать пока рано.Какой бы ни была природа сходства генофондов Северо-Западной России и Северо-Восточной Польши, факт этого сходства генетикой выявлен и даёт возможность верифицировать уже имеющиеся в исторической науке теории появления русского народа и русской государственности.
Сходные популяции: Сувалки (wiki), Великий Новгород, ПсковК процитированному должен добавить следующее. Предшествующие исследования генофондов восточно-европейских народов выявили значительное сходство между поляками, белорусами и русскими в целом — местами даже более значительное, чем между русскими и украинцами. Вместе с этим, по отцовским линиям Y-хромосомы белорусы обнаружили близость не только с поляками, но и с далёкими лужицкими сербами [Балановский, Тегако, 2008]. Такое положение вещей хорошо иллюстрируют карты расстояний на основании данных по Y-хромосоме: от русских популяций, от белорусов и украинцев [Балановский, Балановская, 2008].По результатам автор касается истории и лингвистики, где находит параллель в отмеченных рядом историков следах колонизации областей Северо-Западной России большой группой балтийских славян — выходцев из Южной Прибалтики и Польского Поморья — в 5-6 вв. н.э.При этом речь идёт именно о появлении большой массы славянского населения в Приильменье и верховьях Западной Двины, которое констатируется и археологами, и лингвистами, и антропологами. В то же время, распространённой точкой зрения на появление славян на юго-западном побережье Балтики — предположительно, исходном ареале миграции — является мнение о вытеснении оттуда германцев около 4-5 вв. н.э. и появлении достоверно славянской суковско-дзедзицкой (wiki) археологической культуры. По всей вероятности, вытеснившая германцев группа славян была настолько многочисленна, что буквально за несколько столетий заселила не только свободившиеся земли, но и — морским путём — северо-западные области будущей Руси. Без теории пассионарности за авторством Л.Н.Гумилёва здесь обойтись не удастся.Если абстрагироваться от поиска исторических параллелей и вернуться к популяционной генетике, то указанный сценарий является далеко не единственным. Нам достоверно известен только факт сходства генофондов поляков, русских и белорусов, а также существование внутри них примечательной группы популяций Сувалки-Псков-Новгород, чего самого по себе недостаточно для заключения об имевшей место миграции с запада на восток или с востока на запад. Не менее вероятным выглядит «автохтонный» сценарий, в соответствии с которым указанная группа популяций некогда находилась в пределах единого ареала и была разделена лишь позже. За отсутствием откровенно противоречивых фактов возможность подобного процесса отвергать пока рано.Какой бы ни была природа сходства генофондов Северо-Западной России и Северо-Восточной Польши, факт этого сходства генетикой выявлен и даёт возможность верифицировать уже имеющиеся в исторической науке теории появления русского народа и русской государственности.
Сходные популяции: Сувалки (wiki), Великий Новгород, ПсковК процитированному должен добавить следующее. Предшествующие исследования генофондов восточно-европейских народов выявили значительное сходство между поляками, белорусами и русскими в целом — местами даже более значительное, чем между русскими и украинцами. Вместе с этим, по отцовским линиям Y-хромосомы белорусы обнаружили близость не только с поляками, но и с далёкими лужицкими сербами [Балановский, Тегако, 2008]. Такое положение вещей хорошо иллюстрируют карты расстояний на основании данных по Y-хромосоме: от русских популяций, от белорусов и украинцев [Балановский, Балановская, 2008].По результатам автор касается истории и лингвистики, где находит параллель в отмеченных рядом историков следах колонизации областей Северо-Западной России большой группой балтийских славян — выходцев из Южной Прибалтики и Польского Поморья — в 5-6 вв. н.э.При этом речь идёт именно о появлении большой массы славянского населения в Приильменье и верховьях Западной Двины, которое констатируется и археологами, и лингвистами, и антропологами. В то же время, распространённой точкой зрения на появление славян на юго-западном побережье Балтики — предположительно, исходном ареале миграции — является мнение о вытеснении оттуда германцев около 4-5 вв. н.э. и появлении достоверно славянской суковско-дзедзицкой (wiki) археологической культуры. По всей вероятности, вытеснившая германцев группа славян была настолько многочисленна, что буквально за несколько столетий заселила не только свободившиеся земли, но и — морским путём — северо-западные области будущей Руси. Без теории пассионарности за авторством Л.Н.Гумилёва здесь обойтись не удастся.Если абстрагироваться от поиска исторических параллелей и вернуться к популяционной генетике, то указанный сценарий является далеко не единственным. Нам достоверно известен только факт сходства генофондов поляков, русских и белорусов, а также существование внутри них примечательной группы популяций Сувалки-Псков-Новгород, чего самого по себе недостаточно для заключения об имевшей место миграции с запада на восток или с востока на запад. Не менее вероятным выглядит «автохтонный» сценарий, в соответствии с которым указанная группа популяций некогда находилась в пределах единого ареала и была разделена лишь позже. За отсутствием откровенно противоречивых фактов возможность подобного процесса отвергать пока рано.Какой бы ни была природа сходства генофондов Северо-Западной России и Северо-Восточной Польши, факт этого сходства генетикой выявлен и даёт возможность верифицировать уже имеющиеся в исторической науке теории появления русского народа и русской государственности.{kind=link}
{kind=link}
Происхождение гаплогруппы I1-М253 в Восточной Европе
Александр Штрунов
Происхождение гаплогруппы I1-М253
в Восточной Европе
Абстракт
В современной генетической генеалогии одним из остро обсуждаемых вопросов является вопрос о происхождении той или иной гаплогруппы. Увязать данные генетики, археологии,лингвистики, антропологии и других смежных наук совсем непросто. В этой работе автор попробует найти корни гаплогруппы I1-М253 в Восточной Европе.
Гаплогруппа I1* вместе со своими братскими субкладами I2a* и I2b* является коренной европейской гаплогруппой, частоты гаплогруппы I-M170 за пределами Европы крайне малы. Такое распространение дало основание причислить носителей данной гаплогруппы к палеоевропейскому населению Европы. Ареал гаплогруппы I1-M253 в основном сосредоточен на севере Европы, в Скандинавских странах. Так же существуют локальные максимумы на территории Англии (I1 - 15,4%) [1], Сицилии (I1 - до 18,75%) [2] и в центральной части европейской России (I1 - до 17%) [3]. Присутствие носителей гаплогруппы I1-M253 на Британских островах связывают с экспансиями викингов и норманнов, что подтверждается историческими и генеалогическими исследованиями, хотя и не исключены древние миграции. В Сицилии гаплогруппа I1-M253 достаточно уверенно соотносится с норманнскими походами с территории современной Франции (Нормандии, I1 – 11,9%) и созданием в 1130 году Сицилийского королевства. Однако присутствие гаплогруппы I1-M253 с такими частотами в центре евро- пейской части России вызывает множество вопросов, вот что об этом пишут Е.В. Балановская и О.П. Балановский в своей книге «Русский генофонд на русской равнине»
[4]:
«Генетический рельеф «Скандинавской» гаплогруппы I1a в русском ареале оказывается неожиданным (рис. 1). Казалось бы, высокие значения I1a должны быть вблизи Скандинавии, на северо-западе русского ареала. Там, а также на западной границе можно ожидать «варяжское» влияние в виде повышенных частот I1a. Однако компактный локальный максимум I1a (11-12%) вырисовывается совсем в другой области, на северо-востоке. Это ядро ясно выделяется на фоне окружающих низких частот (менее 6%), свойственных всему остальному русскому ареалу. Реальность этого ядра повышенных значений опирается на данные по трем русским популяциям, изученным по обширным выборкам. Конечно по сравнению с частотами в Скандинавии (25-40%) этот локальный максимум второстепенен. Но его удаленность от основной зоны высоких частот гаплогруппы в Скандинавии требует объяснений. Трудно предположить тесные связи Скандинавии именно с Заволжьем и бассейном Вычегды, помимо прочих русских территорий. Видимо, популяционная история гаплогруппы I1a сложнее, чем простая экспансия из Скандинавии, и, возможно, включает древние связи между финно - угорскими племенами Восточной Европы и предками германоязычных скандинавов».
Рис. 1. Карта распространения гаплогруппы I1 из работы Balanovsky et al. 2008 [5].
Напомним, где наблюдались относительно высокие частоты I1-M253 в работе Balanovsky et al. 2008:
Красноборск, Архангельская область
|
12,1%
|
Вологда
|
11,6%
|
Унжа, Костромская область
|
11,5%
|
Дополним данные по интересующему нас региону при помощи работ других исследователей, для того, чтобы оценить территориальное распространение гаплогруппы I1.
Вологодская область
|
17,0% Roewer et al. 2008 [3]*
|
Архангельск
|
14,2% Mirabal et al. 2009 [6]
|
Рязанская область
|
14,0% Roewer et al. 2008
|
Татарстан
|
13,0% Генофонд.ру**
|
Мокша Старо-Шайгинского района Мордовии
|
12,0 % Rootsi et al. 2004 [7]
|
Пензенская область
|
12,0% Roewer et al. 2008
|
Кострома
|
11,3% Underhill et al. 2007 [1]
|
Тамбовская область
|
10,0% Roewer et al. 2008
|
Ивановская область
|
10,0% Roewer et al. 2008
|
(*определение частот гаплогруппы I1 осуществлялось с помощью предиктора, **данные взяты из атласа на сайте Генофонд.ру)
Данные из дополнительных источников, подтверждают наличие локального максимума I1-M253 на территории центральной части европейской России. На карте ниже
(рис. 2), построенной автором, хорошо видно границы этого локального максимума.
Поскольку подробных исследований по выяснению природы таких высоких локальных частот гаплогруппы I1 со столь обширным ареалом не проводилось, попробуем устранить этот пробел, путем анализа известных источников, увязывая данные генетики, археологии, лингвистики, антропологии и других смежных наук.
Достоверных данных о миграциях из района Скандинавии в историческое время, способных оставить такой след на территории современной России мы не находим, но попробуем рассмотреть все имеющиеся варианты.
Готы Германариха. Германарих или Эрманарих (ум. ок. 375 г) — король готов из рода Амалов. О Германарихе писал готский историк Иордан [8]: «После того как король готов Геберих отошел от дел человеческих, через некоторое время наследовал королевство Германарих, благороднейший из Амалов, который покорил много весьма воинственных северных племен и заставил их повиноваться своим законам. Немало Древних писателей сравнивали его по достоинству с Александром Великим. Покорил же он: гольтескифов, тиудов, инаунксов, васинабронков, меренс, морденс, имнискаров, рогов, тадзанс, атаул, навего, бубегенов, колдов» (здесь исследователи видят будущую формулу русских летописей - Русь, чюдь и вси языци: весь, меря, мордва (?)...)
Германарих и его готы, скорее всего, не могли оставить сколь значимый след на территории современной России и тем более в рассматриваемом нами регионе, поскольку существуют сомнения в существовании столь обширной империи (рис. 3), да и сама империя, если существовала, то век ее был недолог из-за нашествия гуннов. Самих готов связывают с Черняховской археологической культурой (III век н.э.), поэтому следы готов, скорее всего, надо искать среди горной части крымских татар (тата), а также греков Приазовья, сравнивая их гаплотипы с гаплотипами населения севера Испании и острова Готланд.
Варяги. Походы варягов (IX—XII вв.), видимо, тоже могли оставить свой след в генофонде Восточной Европы, однако основные пути варягов проходили в стороне от обозначенных нами территорий. Но и те находки, которые относят к скандинавам, скорее говорят о торгово-экономических отношениях, нежели об экспансии. По этой причине варягов приходится исключить из списка возможных претендентов, которые могли оставить весомый вклад в генофонде Восточной Европы, хотя вероятность того, что какая-то часть варягов могла осесть и ассимилироваться с местным населением, не исключается [10].
Рис. 3. «Империя Германариха» IV в. н.э. и предполагаемый итинерарий III-IV вв. н. э. (схема по Б.А. Рыбакову). а - народы, упомянутые в перечне Иордана; б - порядок перечисления народов; в - основные районы черняховской культуры II-IV вв.; г - направления морских походов III в.; д - направление славянской колонизации III-IV вв. [9].
Древние миграции. Возможно, мы имеем дело с древнейшими миграциями на территории Восточной Европы, о которых предполагали Балановские, что учитывая исключительно европейский ареал гаплогруппы I-M170, представляется разумным.
Если гаплогруппа I1-M253 увязывается с древнейшим населением Европы, то логично предположить, её носители говорили на палеоевропейском языке.
Исследования Серебренникова Б.А. [11] показали, что в междуречье Волги и Оки присутствует древнейшая субстратная топонимия неизвестного происхождения (т.е. неуральская и неиндоевропейская), также она распространена в Нижегородской области (н/д), в Чувашии (I1 - 7,5%), в Кировской (н/д), Вологодской (I1 - 17%), Архангельской областях (I1 - 14,2%), в Карелии(I1 - 8,6%) и на западе в Смоленской области (I1 – 2%).
Примеры субстратной топонимии: на – га (Юзга – пр. Мокши, Арга – пр. Алатыря, Вяжга – пр. Мокши, Волга); на – та (Пущта – пр. Сатиса); на – ша (Кша – пр. Суры, Шокша); на –ма (Лосьма – пр. Мокши, Шалма – пр. Сивини); на - да (Аморда) [12].
Третьяков П.Н. на основании работы Серебренникова выдвинул гипотезу о том, что данный палеоевропейский язык принадлежит создателям, неолитических культур с ямочно-гребенчатой керамикой (ЯГК) [13].
Культура ЯГК первоначально занимала территорию Волго-Окско-Клязьминского междуречья, в III тысячелетии до н.э. ее носители совершают движение на север и северо-запад, где занимают территории от Балтики до Вычегды и Печеры.
Краниологические данные говорят о том, что носители культуры ЯГК льяловского типа крайне неоднородны. В сложении льяловцев участвовало пришлое с севера население лапоноидного облика (рис. 4, справа), и аборигенное европеоидное население волго-окского мезолита пост-свидерской традиции в виде носителей верхневолжской культуры [14].
Рис. 4. Скульптурные реконструкции по черепу мужчины из Володар* (низовья Оки) [15] и по черепу молодого мужчины из погребения № 19 могильника Сахтыш II (Ивановская область) [16].
Верхневолжская культура распространяется в центре Русской равнины на рубеже VI и V тыс., до конца V тыс. до н.э. Ранние верхневолжские памятники сосредоточены ввосточной части своего ареала, а более поздние – в центральной и западной части, что видимо, связано с приходом нового пришлого населения.
Рис. 5. Ареал верхневолжской, волосовской и фатьяновской культур [17].
_________________________
(*данная стоянка относится к более поздней по отношению к верхневолжской волосовской археологической культуре, однако эта скульптурная реконструкция наиболее точно отражает облик европеоидного населения волго-окского мезолита пост-свидерской традиции).
На данный момент версия о происхождении волго-окской культуры с ЯГК от волго-окского мезолита пост-свидерской традиции является наиболее правдоподобной, поскольку переход от мезолита к неолиту в регионе носил плавный характер, и в тоже время бутовская культура, господствующая в конце мезолита в волго-окском междуречье, являлась наследницей свидерских традиций [18].
Свидерская культура – археологическая культура финального палеолита на территории Центральной и Восточной Европы. В связи с изменениями климатических условий в XI – X тыс. до н.э. свидерцы начали движение из района современных Польши, Белоруссии и Литвы на восток и достигли интересующего нас волго-окского региона в VIII тыс. до н.э.
Те же природные процессы заставили мигрировать население носителей соседней и видимо родственной свидерцам аренсбургской традиции. Эта палеолитическая культура существовала в X-IX тысячелетиях до н.э. на территории Дании и Северной Германии, основным занятием ее населения была охота на северного оленя.
В X тысячелетии до н.э. аренсбургцы начали двигаться вслед за отступающим ледником в двух направлениях - на северо-запад и северо-восток, обходя с двух сторон Балтийское ледниковое озеро (рис. 6).
Рис. 6. Этапы в формировании бассейна Балтийского моря [19].
Аренсбургцами был создан ряд так называемых «культур круга маглемозе», которые датируются VIII-VI тыс. до н.э. Это такие культуры, как фосна в Швеции и Норвегии, комса на крайнем севере Скандинавии, включая Кольский полуостров, аскола и суомусъярви в Финляндии и Карелии, веретье в Восточном Приладожье, кунда на территории Приневья, Эстонии и Латвии и маглемозе в Англии, Северной Германии и Дании.
Стоит отметить тот факт, что самое высокое гаплотипное(микросателлитное) разнообразие гапологруппы I1-M253 зафиксировано именно в Дании [1], где находится отправная точка аренсбургцев.
Рис. 7. Истоки палеоевропейских субстратных компонентов в палео- и мезолите [18].
Условные обозначения: 1 – первоначальный ареал и распространение аренсбургской культуры; 2 – первоначальный ареал и распространение свидерской культуры; 3 – район формирования культур ЯГК волго-окского типа.
Если предположения о родстве носителей аренсбургской и свидерской культур верны, то мы должны наблюдать на территории Скандинавии схожую антропологическую, лингвистическую и генетическую картину.
Антропологически носители аренсбургской традиции имели характерный резко долихокранный (рис.8.), широколицый, высокорослый европеоидный тип, который вполне соответствует пост-свидерскому аборигенному типу в волго-окском междуречье, имевшему ярко выраженный европеоидный облик, долихокранный со среднешироким лицом. Тот же антропологический тип участвовал в сложении саамов. Сопоставление данных антропологов показывает, что в лопарском типе присутствуют черты древнего североевропеоидного населения и более поздние черты, связанные с проникновением на север монголоидного комплекса. Слияние двух компонентов и привело к формированию антропологических особенностей лопарей. В одонтологии также выделяют наличие лопарей древнего северного реликтового и более позднего монголоидного комплекса [20].
Рис. 8. Примеры различных форм черепа [21].
В саамском языке так же выделяют два генетических компонента, первый – субстратный дофинский, второй – древнефинский, близкий к прибалто-финским и волго-финским языкам. Наследие первого особенно хорошо прослеживается в лексике, в меньшей мере в морфологии, фонетике и синтаксисе. По подсчетам специалистов, до одной трети саамской лексики имеет субстратный характер, которая не имеет аналогий ни в одном из существующих языков мира [20].
Генетические данные говорят о том, что гаплогруппа I1, с которой мы связываем палеоевропейское население Северной Европы, в генофонде саамов имеет следующие частоты: саами-28% [7], шведские саамы - 32% [22], анара-саами-34% [23]*, скола-саами-52% [23], саами ловоозерские-17% [23].
На основании выше приведенных данных гипотеза о связи гаплогруппы I1 и палеолитического населения Европы выглядит крайне убедительной.
Анализ источников позволяет реконструировать популяционную историю гапло- группы I1-M253, таким образом:
Как и когда носители гаплогруппы I оказались в Европе - пока не ясно, и этот вопрос сейчас является дискуссионным, также не ясно, когда и где I1 отделилось от I. Однако в палеолите носители гаплогруппы I1 оказались в центральной Европе в северной ее части на территории нынешних Дании, Северной Германии и Польши. Они создали такие культуры, как аренсбургская и свидерская (автор не той исключает возможности, что в сложении свидерской культуры участвовали и носители гаплогруппы I2a), основным их занятием являлась охота (преимущественно на северного оленя, лося и бобра) и собирательство.
_________________________
(* гаплогруппа I1 в данной работе не типирована, но результаты других работ [24] и публичных ДНК проектов [25] позволяют с уверенностью об этом заявлять)
В XI – X тыс. до н.э. происходили глобальные климатические изменения, которые привели к таянью ледникового щита в Скандинавии. На освободившиеся от ледника территории проникала растительность, вслед за которой двигался и основной источник пищи людей этого региона – северный олень, что заставило двигаться и палеолитических охотников. Осваиваются Скандинавия, Прибалтика и центральная часть Восточной Европы.
В мезолите носители гаплогруппы I1 столкнулись с восточными пришельцами, которые уверенно соотносятся с гаплогруппой N1c, они имели уралоидный облик (сочетающий в себе как монголоидные, так и европеоидные черты) и именно с ними связано распространение уральских языков на территории Восточной Европы.
Отношения между пришельцами и местным населением в целом были мирные, это видно по смешанным захоронениям и постепенному появлению метисных антропологических типов.
Следующий заметный штрих в формировании существующей картины внесли носители культур со шнуровой керамикой, у которых обнаружена гаплогруппа R1a [27] и которых следует связывать с распространением в Средней Европе южноевропейских земледельческих и скотоводческих племен.
В III тыс. до н.э. «шнуровики» из Центральной Европы проникли в Прибалтику (прибалтийская культура боевых топоров) и в районы верхней и средней Волги (фатьяновская (Рис.5.) и балановская культуры).
Антропологически они представлены резко долихокранным, умеренно широколицым европеоидным типом [28].
По всей вероятности, «шнуровики» были достаточно агрессивными, они вытеснили в наиболее глухие районы и частично ассимилировали местное население (I1/N1c), которое не могло конкурировать с пришельцами в экономическом и в военном плане, так как не было знакомо с металлургией и производящим хозяйством [29].
Так же следует отметить достаточно массовые миграции славянских племен (VII-VIII вв.), которые внесли заметные изменения в генофонд Восточной Европы. В основной массе славяне являлись носителями гаплогруппы R1a (в большей степени) и I2a.
В результате этих процессов носители гаплогруппы I1 были частично вытеснены из своих ареалов обитания, а частично ассимилированы более развитыми пришельцами.
В заключении хотелось бы выделить основные выводы работы:
- Корни гаплогруппы I1 достаточно убедительно находятся в таких палеолитических культурах, как аренсбургская и свидерская, ее носители являлись автохтонным населением Северной и Восточной Европы.
- Основным видом деятельности носителей гаплогруппы I1 являлась охота и собирательство.
- Первоначальный антропологический облик носителей гаплогруппы I1 выражен резко долихокранным, широколицым, высокорослым европеоидным типом.
- Носители гаплогруппы I1 обладали собственным палеоевропейским языком не принадлежавшим уральским и индоевропейским семьям, который обнаруживается в европейской топонимике и языке саамов.
- Компактный локальный максимум частот I1 в центре Русской равнины, - следствие древнейших миграций палеолитического населения Европы, приведших к созданию верхневолжской культуры (VI – V тыс. до н.э.).
Литература
1. Underhill PA, Myres NM, Rootsi S et al: New phyloge netic relationships for Y-chromosome haplogroup I: reappraising its phylogeography and prehistory; in Mellars P, Boyle K, Bar-Yosef O, Stringer C (eds): Rethinking the Human Revolution. Cambridge, UK: McDonald Institute Monographs, 2007, pp 33– 42.
2. Di Gaetano C, Cerutti N, Crobu F et al: Differential Greek and northern African migrations to Sicily are supported by genetic evidence from the Y chromosome. Eur J Hum Genet 2009; 17: 91– 99.
3. Roewer L., Willuweit S., Krüger C. et al. 2008 Analysis of Y chromosome STR haplotypes in the European part of Russia reveals high diversities but nonsignificant genetic distances between populations Int J Legal Med.
4. Балановская Е. В., Балановский О. П. Русский генофонд на Русской равнине. – М.: Луч, 2007.
5. Balanovsky et al. Two sources of the Russian patrilineal heritage in their Eurasian context. Am J of Hum Genet 2008; 82: 236– 250.
6. Mirabal S, Regueiro M, Cadenas AM et al. (2009) Y- Chromosome distribution within the geo-linguistic landscape of northwestern Russia. Eur.J.Hum.Genet.
7. Rootsi et al. 2004 Phylogeography of Y-Chromosome Haplogroup I Reveals Distinct Domains of Prehistoric Gene Flow in Europe, Am. J. Hum. Genet. , 2004, vol. 75, pp. 128–137.
8. Иордан. О происхождении и деяниях гетов. Спб. Алетейя. 1997.
9. Рыбаков Б.А. - Язычество Древней Руси.1987.
10. Пушкина Т.А. О проникновении некоторых украшений скандинавского происхождения на территорию
Древней Руси//Вестник МГУ. История. 1972. № 1.
11. Серебренников Б. А. 1955. Волго-окская топонимика на территории Европейской части СССР // Вопросы языкознания. №6. Москва.
12. Цыганкин Д.В. Мордовская архаическая лексика в топонимии Мордовской АССР. / «Ономастика Поволжья». Выпуск 4. – Саранск, 1976.
13. Третьяков П. Н. 1958. Волго-окская топонимика и вопросы этногенеза финно-угорских народов // Советская этнография. №4. Москва.
14. Костылева Е.Л., Уткин А.В. Краткая характеристика антропологических типов эпохи первобытности на территории Ивановской области, Проблемы отечественной и зарубежной истории: Тез. докл. Иваново, 1998. С. 55-57.
15. Герасимов М. М. Восстановление лица по черепу: современный и ископаемый человек. - М., 1955. - С. 377.
16. Лебединская Г.В. Облик далеких предков: Альбом скульптурных и графических изображений. - М.: Наука, 2006.
17. Рисунок взят с сайта http://www.prezidentpress.ru/ 18. Напольских В.В. Происхождение субстратных палеоевропейских компонентов в составе западных финно-угров. // Балто-славянские исследования 1988-1996. М., 1997.
19. Рисунок взят с сайта Георгия Хохлова http://hohloff-spb.narod.ru/cayt/golocen.html
20. Манюхин И.С. Этногенез саамов Ижевск 2005 (опыт комплексного исследования). http://elibrary.udsu.ru/xmlui/bitstream/handle/12345678 9/655/05_12_010.pdf?sequence=2
21. Рисунок взят с сайта http://www.balto-slavica.com/ Балто-Славика,
22. Karlsson, A. O., Wallerstrom, T., Gotherstrom, A., Holmlund, G. (2006) Y-chromosome diversity in Sweden – a long-time perspective. Eur. J. Hum. Genet. 14, 963–70.
23. Raitio M., et al., Y-chromosomal SNPs in Finno-Ugricspeaking populations analyzed by minisequencing on
microarrays, Genome Res. 11 (2001) 471–482.
24, Pericic, M., Lauc, L.B., Klaric, A.M. et al. High-resolution phylogenetic analysis of southeastern Europe traces major episodes of paternal gene flow among Slavic populations. Mol. Biol. Evol. 22, 1964-1975 (2005).
25, Family Tree DNA Saami Project http://www.familytreedna.com/public/Saami/default.aspx
26 Рисунок взят с сайта http://oldtiden.natmus.dk/ Haak, W., Brandt, G., de Jong, H., Meyer, C., Ganslmeier, R., Heyd, V., Hawkesworth, C., Pike, A., Meller, H., and Alt, K. (2008). Ancient DNA, Strontium isotopes and osteological analyses shed light on social and kinship organization of the Later Stone Age. Pro-ceedings of the National Academy of Sciences of the United States of America 105, 18226-18231.
28,Денисова Р.Я. 1975. Антропология древних балтов. Рига.
29,Крайнов Д.А., Лозе И.А., 1987. Культуры шнуровой керамики и ладьевидных топоров в Восточной Прибалтике. // Археология СССР. Эпоха бронзы лесной полосы СССР. М.
30, Бадер О.Н. О древнейших финно-yrpax на Урале и древних финнах между Уралом и Балтикой // Проблемы археологии и древней истории угров. М., 1972.
31, Боготова З.И. Изучение генетической структуры популяций кабардинцев и балкарцев. Уфа – 2009 Автореферат диссертации на соискание ученой степени кандидата биологических наук.
32, Лобов А.С. Структура генофонда субпопуляций башкир. Автореферат на соискание ученой степени кандидата биологических наук. Уфа. 2009. С.15.
33, Напольских В.В. К реконструкции лингвистической карты Центра Европейской России в раннем железном веке // Арт. №4. Сыктывкар, 2007; стр. 88-127.
34, Напольских В.В. Предыстория уральских народов // Acta Ethnographica Hungarica. T. 44:3-4. Budapest, 1999; стр. 431-472.
35,Харьков В.Н., Степанов В.А., Медведева О.Ф. и др. Различия структуры генофондов северных и южных алтайцев по гаплогруппам Y-хромосомы // Генетика. 2007. Т. 43. № 5. С. 675–687.
36, Харьков В.Н., Медведева О.Ф., Лузина Ф.А., Колбаско А.В., Гафаров Н.И., Пузырев В.П., СтепановВ.А.. Сравнительная характеристика генофонда телеутов по данным маркеров Y-хромосомы. Генетика. 2009. Т. 4.
40.Юнусбаев Б.Б.. Популяционно-генетическое исследование народов Дагестана по данным о полиморфизме Y-хромосомы и Alu-инсерций: дис. канд. биол. наук: 03.00.15 Уфа, 2006 107 с. РГБ ОД, 61:07-3/183.
41,Abu-Amero et al Saudi Arabian Y-Chromosome diver-sity and its relationship with nearby regions // BMC Genetics 2009, 10:59 doi:10.1186/1471-2156-10-59.
АНТРОПОЛОГИЯ ДОНСКИХ КАЗАКОВ: ОПЫТ
ИНТЕГРАЦИИ ДАННЫХ НАУКИ И ЛИТЕРАТУРЫ
В.Ф.
Кашибадзе, О.Г. Насонова
Антропологическая история донских казаков
подразумевает процессы миграции из юго-восточных зон Центральной
России и незначительное включение южных и восточных элементов в возрастающей к
югу пропорции. Совпадение антропологических данных начала и второй половины ХХ
века указывает на преемственность физических характеристик этой группы
населения и, отсюда, на выживание группы в процессе
известных демографических коллизий. Статистический анализ антропологического
портрета донских казаков в романе-энциклопедии М.А. Шолохова «Тихий Дон»
показал соответствие портрета научным данным и воспроизведение в романе достоверной антропологической среды.
ANTHROPOLOGY
OF THE DON COSSACKS: EXPERIENCE BY INTEGRATION OF
SCIENTIFIC
AND LITERARY DATA
V.F.
Kashibadze, O.G. Nasonova
The anthropological history of the Don Cossacks implies their
ancestors' expansion from the mentioned areas to the South and negligible
influence of southern and eastern elements in the southward increasing ratio.
The correspondence of the data obtained in the 10th and
in the 70th years of the 20th century provides the evidence of continuity in physical
characteristics of the group and of its survival in the process of undergone
demographical collisions. The statistical textual analysis of the Don Cossacks anthropological portrait in M. Sholokhov novel-encyclopedia The
Quiet Don reveals the accordance of the portrait to the scientific data proving
the reproduction of the authentic anthropological environment in the novel.
Целью
настоящего исследования является представление одонтологической
характеристики одной из наиболее исторически и социально значимых групп
населения южного региона - донских казаков - на основе многомерного
статистического анализа оригинальных современных и ископаемых материалов. Кроме
того, в работе сделана попытка обобщить всю имеющуюся информацию по
антропологии этой группы, взятую как из научных, так и из художественных
источников.
Основными подходами послужили
принципы антропофенетики, сформулированные в последние
десятилетия [12, с. 9-11, 13, 15, с. 21-22, 17, с. 16-30, 22, с. 7-15], а также
использование созданной авторами реляционной базы данных по одонтологии
евразийских популяций. Отдельной отправной позицией является установленное
междисциплинарными исследованиями определение романа М.А.
Шолохова «Тихий Дон» [23] как энциклопедии донского казачества. Исследователями
творческого наследия писателя отмечена феноменальная точность автора при
использовании им письменных источников, устных свидетельств, своих собственных наблюдений [8] в определении хронологии событий, в описании
отношений людей, быта казачества, структуры казачьих войск, топонимических,
демографических и агрономических характеристик, семейного быта казаков,
реальных событий исторического значения, пейзажей, растительности [20],
[21], астрономического ландшафта [16].
Материалами для выполнения работы
послужили одонтологические данные по 2 выборкам: современная популяция
верхнедонских казаков из станицы Вешенской численностью 181 чел., обследованная нами в 1976 г., и краниологическая серия нижнедонских казаков XVIII
в. численностью 67 индивидуумов из коллекций лаборатории физической
антропологии ЮФУ-ЮНЦ РАН, изученная нами в 2008 г. Сравнительное пространство
составили данные по 442 выборкам современного населения Евразии, относимого
к западным (европеоидным) формам, причем 114 из них были изучены авторами,
данные по остальным взяты из научной печати. Другой базовой составляющей
материалов являются результаты произведенного нами текстологического анализа антропологического портрета в романе М.А. Шолохова «Тихий Дон»
[23]. Сравнительное пространство в этой части составили данные важнейшего
исследования антропологии донских казаков, осуществленного одним из
основоположников отечественной науки о человеке
академиком В.В. Бунаком в 1912-1915 гг.
Одонтологический материал был собран и
исследован по методике и программе, разработанной отечественной научной школой
[9, 10, 11, 12, 13]. Статистический анализ одонтологических данных проводился
методом главных компонент с помощью алгоритмов, написанных
на языке python. На нем же были реализованы и
графические построения.
Одонтологическое
изучение донских казаков было начато еще в 1976 г. в рамках большого
исследования русских Европейской части России [5, 6, 7]. Напомним, что признаки зубной системы человека являются максимально
информативными для решения задач исторического характера. Одонтологией к
настоящему времени накоплен огромный массив
данных, представляющих временные
и пространственные характеристики
Впервые на большом материале,
представляющем различные территориальные группы Европейской части России, была
дана антропологическая характеристика русских по системе одонтологических
признаков: установлено, что одонтологический комплекс русских составляют
компоненты двух основных типов - среднеевропейского и северного грацильного;
впервые на большом пространственном расстоянии прослежено взаимодействие этих двух типов через ряд переходных форм, обусловленных наличием
непрерывной и направленной изменчивости дифференцирующих признаков, что
подтверждает реальность существования выделенных одонтологических типов и
созданной классификации в целом; получены новые
данные о распределении далеко на Юге, вплоть до центральных и черноземных зон
Европейской части России, отдельных вариантов северного грацильного типа, по
форме отличающихся от уже известных, что позволяет расширить представления о
северном грацильном типе и его природе; установлено, что
при наличии определенного сходства с соседними народами Восточной Европы
русские формируют отчетливое единство по системе одонтологических признаков
[7,
с. 4-5].
Данные по наиболее важным
дифференцирующим признакам в русских и сравнительных популяциях
представлены в таблице 1.
Наиболее
общие и визуализированные, в силу огромного массива данных, результаты проведенного в масштабе суммарных этнических групп как систем
популяций, обладающих генетической памятью, компонентного анализа европеоидных
групп Евразии [15, с. 149] определяют положение русских между северными
грацильными и среднеевропейскими формами включительно (рис. 1).
Среди изученных 27 групп русского
населения самой репрезентативной была выборка из станицы Вешенской. И в
общерусском масштабе, и на фоне европеоидных групп Евразии она показывает
поразительное сходство с группой из Рязанской области.
По ряду признаков (диастема, редукция верхнего латерального резца,
одонтоглифические признаки-фены) такое сходство принимает качество тождества,
тем более выразительного, что население соседних областей (Воронежской,
Липецкой, Тамбовской, Белгородской)
В
июле 2008 года авторами была изучена репрезентативная краниологическая серия
казаков XVIII в. Нижнего Дона,
добытая в ходе археологических раскопок, в основном, на территории современного
города Ростова-на-Дону, из коллекций лаборатории физической антропологии
ЮФУ-ЮНЦ РАН. Анализ одонтологических характеристик этой группы позволил сделать
заключение о том, что по основному набору признаков она
ближе всего к выборке современных донских казаков из станицы Вешенской и к
современным русским Рязанской области. Более того, по таким важным
диагностическим маркерам, как лопатообразные формы верхних резцов и вариант 2med II,
казаки XVIII в. оказываются даже ближе к рязанцам, чем вешенцы. Отличается эта
группа нижних донцов от рязанцев и, тем более, от других русских групп в том же
направлении, что и вешенцы от русских Центральной России: при сохранении
одонтологического базиса последних, отмечается слабое
присутствие восточных и неких южных элементов, вполне естественных для ранних
мигрантов в степную зону.
Данные исследования географии
русских фамилий свидетельствуют о наличии интенсивных миграционных процессов в XV-XVIII в.
из центра на юг, связанных с освоением новых необжитых окраин «Дикого поля».
Например, распространение фамилий Попов, Богатырев, характерных для
верхнедонских казаков, фактически совпадает с границами Русского государства
этой эпохи и маркирует миграционные процессы на
юго-восток, по берегам Дона [1, с. 149, 18].
По данным сравнительного анализа
донором южной компоненты мог быть аланский одонтологический комплекс,
являющийся важным формообразующим фактором и для групп Северного Кавказа [15,
с. 135-136]. Вклад украинской компоненты в одонтологический статус этой
группы нижних донцов полученными данными не фиксируется.
Нами был произведен анализ
одонтологических показателей репрезентативного ряда русских групп (Псковской,
Новгородской, Рязанской, Тамбовской, Липецкой, Воронежской,
Курской и Белгородской областей), обеих выборок казаков и характерных



Значения
коэффициентов главных компонент для совокупности подвергнутых анализу групп
дифференцируют главные одонтологические комплексы популяций Восточной Европы вообще: северный грацильный, южный грацильный и среднеевропейский. В
пространстве этих комплексов взятые для настоящего анализа выборки занимают
соответствующие позиции. Русские образуют широкий, но собственный разброс в
правом поле, причем один из наиболее отчетливых кластеров этого разброса
составляют три объектных группы: русские Рязанской области, верхнедонские
казаки Ростовской области и нижнедонские казаки XVIII в.
(рис. 2). Украинцы, адыгейцы и осетины занимают соответствующие места в левом
поле графика.
Таким образом, главным выводом
проведенного одонтологического исследования донских казаков является заключение
о том, что в основе их физического статуса лежат фенетические характеристики,
общие с русским населением юго-восточных зон Центральной России. Морфологически и генетически ближайшей к ним группой являются русские
Рязанской области.
Важнейшее исследование антропологии
донских казаков было осуществлено одним из основоположников отечественной науки
о человеке академиком В.В. Бунаком [2] в 1912-1915 гг.
Статья В.В. Бунака, несмотря на ее сравнительно небольшой объем, является
энциклопедичным исследованием. Автор приводит данные по истории заселения Дона
и ценные демографические характеристики. Так, по материалам Областного
статистического комитета общая численность донского казачества
в 1914 г. определялась в 1 426 561 чел., что составило 44,5 % населения всей
Донской области, причем в 19101914 гг. происходило относительное
убывание казачьего населения. Распределение казачества по полу дает следующие соотношения: мужчин - 51,4 %, женщин - 49,6 % . В возрастном
распределении значительно преобладает молодежь до 18 лет - 50,8 % юношей
Сам
В.В. Бунак работал в 5 районах, соответствующих основным
историко-этнографическим провинциям Донской области: нижнедонском, донецком,
среднедонском, верхнедонском, хоперском. В каждом районе изучено по 50 человек, всего по области - 250 человек. Программа исследований
включала классические измерительные и описательные признаки головы, лица и
тела: окраска глаз, волос, рост, головные и лицевые указатели.
Полученные В.В. Бунаком результаты
таковы. Донских казаков следует отнести к числу
сравнительно светлых по окраске групп русского населения и сблизить их в этом
признаке не с непосредственными соседями, а с населением более удаленных
районов средней и северной России. Малороссы оказываются более темнопигментированными. То же направление морфологических связей казаков, а
именно со средней Россией, определяют и значения головного указателя [2, с.
139-143]. Донцы - одна из самых высокорослых групп России. По этой особенности
они сближаются также с другими казаками - кубанскими и терскими -
и отличаются от неказачьего населения тех же областей [2, с. 140].
Согласно
Бунаку, для донских казаков характерны следующие особенности: прямые или слегка
волнистые волосы, густая борода, прямой нос, широкий разрез глаз, крупный рот, русые или светло-русые волосы, серые, голубые или
смешанные (с зеленым) глаза, относительно высокий рост и относительно широкое
лицо. По этим характеристикам донцы входят в круг морфологической изменчивости
у русских и отличаются от украинцев. Незаметно также сколько-нибудь
значительных следов примеси инородной крови; они прослеживаются лишь в
отдельных пунктах (в основном, на юге и западе) и в слабом количестве [2, с.
144-145].
В.В. Бунак отмечает, что донские
казаки, тем не менее, имеют ряд особенностей, отличающих их от
других групп русского населения (комбинация светлой окраски, высокорослости,
широколицести и мезоцефалии), что составляет не их исключительную
принадлежность, а общую у них с другими казачьими группами. Вследствие этого, в
приведенных выше отличиях следует видеть не только тип донских казаков,
но и тип казачества вообще [2, с. 145]. Выявленную особенность ученый объясняет
тем, что казачество является населением колонизационного типа, сложившимся
сравнительно
недавно
и до известной степени искусственно образовавшимся, претерпевшим очевидные
процессы смешения между русскими - выходцами из разных областей и районов
России [2, с. 146]. В.В. Бунак скрупулезно рассматривает территориальные вариации признаков в Донской области и делает на основе их анализа
предположения, население какой зоны Великороссии составило основу физического
типа той или иной группы донских казаков [2, с. 147-148].
Итак,
по своим антропологическим характеристикам донские
казаки определяются В.В. Бунаком как: а) часть русского народа; б) группа с
отличительными для казачества вообще особенностями; в) население, имеющее
относительные локальные различия. Результаты антропологических исследований
позволяют сделать самый важный вывод: донские казаки плотью и
кровью являются частью русского народа. В основе их физического статуса лежат
морфологические характеристики, общие с населением юго-восточных зон
Центральной России, указывая тем самым на направления генетических связей. Антропологическая история донских казаков подразумевает процессы
смешения между разными исходными группами русских, возможно, адаптацию к новым
природным и социальным условиям, а также незначительное включение южных и
восточных элементов в возрастающей к югу пропорции.
Наше внимание давно привлек один из
аспектов информационной составляющей произведений М.А. Шолохова, а именно
антропологический портрет. От художественного он отличается точностью
воспроизведения физического облика окружавшего писателя населения - донских казаков. Приводимые в текстах портреты кратки и
выхватывают самое существенное в облике персонажа. Гипотезой предполагаемого
исследования в этом направлении является допущение, что антропологический
портрет в работах М.А. Шолохова воспроизведен так
же точно, как и все другие стороны мира донских казаков, и соответствует их
научным морфологическим характеристикам.
Работа В.В. Бунака по донским
казакам, опубликованная в 1922 г. в «Русском антропологическом журнале»,
представляет большую ценность, поскольку зафиксировала физический
статус казаков начала ХХ века, до революции и гражданской войны, повлекших
значительные демографические изменения в Области войска Донского. Фактически
исследованные В.В. Бунаком казаки и являются по времени действующими лицами романа «Тихий Дон», что делает сопоставление литературных и
научных данных в этом случае совершенно корректным. Сравнение литературной
информации с материалами В.В. Бунака проводится по признакам классической
антропологии, из которых выбраны четыре, которые могут быть корректно с
научной точки зрения отслежены в тексте романа: цвет глаз, цвет волос, цвет
бороды и усов, а также форма
Всего на страницах романа нами было
найдено 133 персонажа, определенных автором как казаки. Выборку такого объема в
антропологической науке считают репрезентативной. Ее особенностью, тем не
менее, является то, что не все из 133 персонажей описаны М.А. Шолоховым по полной выбранной программе. Для большинства из них, а это
второстепенные или эпизодические действующие лица, выхвачена одна и самая
существенная черта в их внешнем облике: голубые глаза, курносый нос, рыжая
борода, пшеничные усы и т.д. Численность наблюдений по отдельным
обозначенным признакам, таким образом, ниже, она и приводится в основной
таблице данных. Тем не менее, статистика по роману вполне сопоставима с таковой
у В.В. Бунака.
Цветность в антропологии определяется по весьма подробным шкалам
Фишера. Для наиболее общих выводов эти шкалы были модифицированы В.В. Бунаком,
классиком отечественной антропологии. Так, из 30
оттенков цвета волос по Фишеру нами рассмотрены три объединенные Бунаком
группы: темные, переходные и светлые. Двенадцать градаций цвета глаз так же
рассматриваются в трех вариантах: темные (черные и карие), переходные
(буро-желто-зеленые, зеленые, серо-зеленые, серые или голубые с
буро-желтым венчиком), светлые (серые, голубые, синие). Форма носа (спинка
носа) бывает прямая, вогнутая (курносый), выпуклая (горбоносый) и
комбинированная. Естественно, М.А. Шолохов не отмечает в портретах своих героев
наиболее распространенные прямые спинки носа, но всегда упоминает курносых
и горбоносых. Все упомянутые посылки и определили методологический подход к
сравнению данных по донским казакам классика отечественной науки и классика
мировой литературы.
Удивительное соответствие обеих групп данных не требует комментариев.
Единственным заметным расхождением является более высокий процент темных глаз
(за счет переходных) у казаков из романа «Тихий Дон». Черноглазыми являются
главные герои романа. Возможно, в данном случае выразились личные
эстетические предпочтения М.А. Шолохова. Другим и не противоречащим первому, на
наш взгляд, будет предположение о намеренном выделении писателем основных
героев - членов семьи Мелеховых и Аксиньи - из окружающей их среды одним штрихом, одним мазком. Очевидно, это та тонкая грань, тот изящный баланс
типичности-необычности, который находят великие художники.
Проведенное исследование
антропологического портрета осветило еще одну сторону энциклопедичности романа
М.А. Шолохова «Тихий Дон». Удивительная точность,
способность видеть правду жизни во всех ее ипостасях и воспроизводить ее кистью
художника создают ощущение присутствия автора во всех событиях произведения и
позволяют читателю погрузиться в достоверную, живую среду мира донских казаков. Статистический анализ антропологического портрета донских
казаков в романе-энциклопедии М.А. Шолохова «Тихий Дон» показал соответствие
портрета научным данным и воспроизведение в романе достоверной
антропологической среды.


Список литературы:
1.
Бужилова А.П. География
русских фамилий // Восточные славяне. М.: Научный мир, 1999. С. 135-151.
2.
Бунак В.В. Антропологический
тип донских казаков // Русский антропологический журнал, М.: 1922. Т. 12. Вып.
1-2. С. 130-154.
3.
Бунак В.В. Областные
типы // Происхождение и этническая история русского народа. Труды Ин-та
этнографии. Новая серия. 1965. Т. 88. С. 152-162.
4.
Бунак В.В. Некоторые
вопросы этнической истории // Происхождение и этническая история русского
народа. Тр Труды Ин-та этнографии. Новая серия. 1965. Т. 88.
С.
256-270.
5.
Ващаева (Кашибадзе) В.Ф. Одонтологическая
характеристика русских западных и северо-западных областей РСФСР // Вопросы
антропологии. Вып. 56. М.: Изд-во
Москов. ун-та, 1977. С. 102-111.
6.
Ващаева (Кашибадзе) В.Ф. Одонтологическая
характеристика русских центральных, южных и северных областей Европейской части
РСФСР // Вопросы антропологии. Вып. 57. М.: Изд-во Москов. ун-та, 1978. С.
133-142.
7.
Ващаева (Кашибадзе) В.Ф. Одонтологическая
характеристика русского населения Европейской части РСФСР: Автореф. дисс.
канд. биол. наук. М., 1978. 20 с.
8.
Ермолаев Г.С. Михаил
Шолохов и его творчество. СПб.: Речь, 2000. 324 с.
9.
Зубов А.А. Одонтология:
методика антропологических исследований. М.:
Наука,
1968.
200 с.
10.
Зубов А.А. Этническая
одонтология. М.: Наука, 1973. 203 с.
11.
Зубов А.А. Одонтоглифика
// Расогенетические процессы в этнической истории. М.: Наука, 1974. С. 11-42.
12.
Зубов А.А., Халдеева Н.И. Одонтология
в современной антропологии. М.: Наука,
1989.
232 с.
13.
Зубов А.А., Халдеева Н.И. Одонтология
в антропофенетике. М.: Наука, 1993. 223 с.
14.
Кашибадзе В.Ф. Об
антропологическом портрете в произведениях М.А. Шолохова // Вешенский вестник.
Ростов-н/Д: Ростиздат, 2005. С. 82-89.
15.
Кашибадзе В.Ф. Кавказ
в антропоисторическом пространстве Евразии. Ростов н/Д.: Изд-во ЮНЦ РАН, 2006.
312 с.
16.
Кашибадзе В.Ф., Кашибадзе О.Г. Астрономический
ландшафт в произведениях М.А. Шолохова. URL. http://www.pereplet.ru:
Русский переплет. Литературный интернет-журнал. М., 2005. URL: http://www.pereplet.ru/text/kashibadze/astroland
(дата обращения 16.05.2005) // Вешенский вестник. Ростов-н/Д:
Ростиздат, 2006. С. 46-54.
17.
Мовсесян А.А. Фенетический
анализ в палеоантропологии. М.: Университетская
книга,
2005. 272 с.
18.
НиконовВ.А. География
фамилий. М.: Наука, 1988. 192 с. С. 23-28.
19.
Сегеда С.П. Украинцы
// Этническая одонтология СССР. М.: Наука, 1979. С.
32-46.
20.
Семанов С.Н. В
мире «Тихого Дона». М.: Современник, 1987. 253 с.
21.
Степаненко Л.Г. Ростовская
область: растительное убранство. Ростов н/Д.: Багир,
2002.
248 с.
22.
Хить Г.Л., Долинова Н.А. Расовая
дифференциация человечества. Дерматоглифические
данные. М.: Наука, 1990. 206 с.
23.
Шолохов М.А. Тихий
Дон. В 4 кн. Ростов н/Д.: Ростовское кн. изд-во, 1978. 1227 с.