Волков В.Г., Харьков В.Н., Степанов В.А. Андроновская и тагарская культуры в свете генетических данных.
//Труды Томского областного краеведческого музея им. М.Б.Шатилова.- Томск, 2012.- Т.XVII.- C.147 - 166.
//Труды Томского областного краеведческого музея им. М.Б.Шатилова.- Томск, 2012.- Т.XVII.- C.147 - 166.
Современные методы, применяемые в молекулярно-генетических исследованиях, позволяют исследовать не только генофонд современного населения, но и древних популяций. Извлекая ДНК из останков носителей древних археологических культур и сравнивая с ДНК современных людей, можно с достаточно большой точностью установить, какие народы являются потомками древней культуры, а также пути миграций древнего населения (Keyser-Tracqui C. et al., 2003; Schilz, 2006; Chikisheva et al., 2007; Gao et al., 2008; Krause et al., 2009; Zhao et al., 2010).
Особое значение в этих методах придается исследованию нерекомбинируемых участков Y-хромосомы. Y-хромосома передается только по мужской линии, и при этом отсутствует какое-либо влияние на нерекомбинируемые участкиY-хромосомы других наследственных факторов, и это позволяет сравнивать между собой разные мужские линии и устанавливать степень родства между ними.
В 2009 году были опубликованы результаты исследований древней ДНК носителей андроновской, тагарской и таштыкской культур (Keyser et al., 2009)
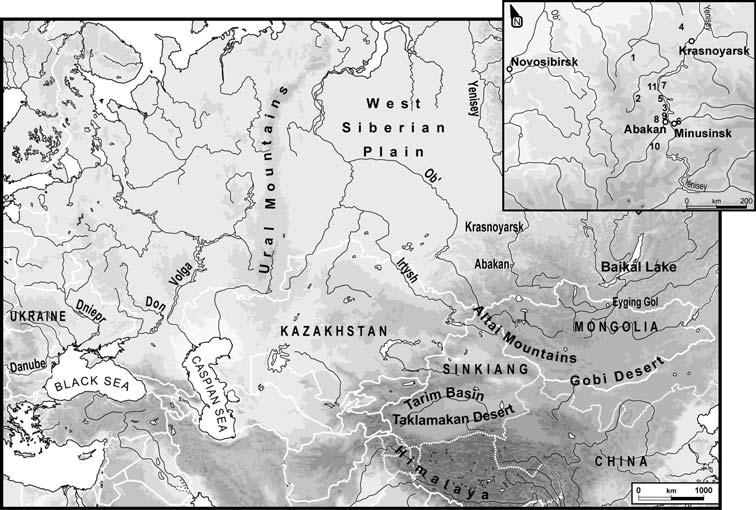
Ил.1. Карта расположения захоронений (из работы Keyser et al., 2009)
Были исследованы образцы из следующих захоронений андроновской культуры:
1. Шарыповский район, Красноярского края - Андроновская культура 1800–1400 вв. до н.э. (S07)
2. Красноярский край - Андроновская культура 1800–1400 вв. до н.э. (S10).
3. Хакассия - Андроновская культура 1800–1400 вв. до н.э. (S16).
Определены были Y-гаплогруппы и Y-STR гаплотипы погребенных в этих захоронениях людей.
Первый гаплотип андроновцев, обозначенный как S07, относится к гаплогруппе C.
ANDRON S07 14-22-15-9-12-13-11-11-12-16-16-14-10-15-19-11-22
[Порядок маркеров: DYS393 DYS390 DYS19 DYS391 DYS385a DYS385b DYS439 DYS389I DYS392 DYS389II DYS458 DYS437 DYS438 DYS456 DYS448 DYSH4 635]
Эта гаплогруппа распространена в основном в Восточной и Юго-Восточной Азии и, соответственно, носитель данного гаплотипа не связан генетически с основной группой андроновцев, а является потомком автохтонного населения.
Самый близкий вариант при различии 5 мутаций на 10 маркерах присутствует у эвенка (Malyarchuk, 2010).
230-C3c-13-24-16-9-12-13-11-11-11-16-17-14-10-15-20-10-23
Также к этому гаплотипу близки гаплотипы тувинцев, монголов и калмыков. Но он не может быть предковым для гаплотипов этих народов, а лишь связан с ними очень отдаленным родством. Мы можем лишь предполагать, что предки носителя этого гаплотипа мигрировали с территории Манчжурии через Прибайкалье или Монголию к Енисею и Чулыму.
Гаплотипы андроновцев, обозначенные как S10 и S16, идентичны и относятся к гаплогруппе R1a1a, определяемой SNP-маркёрами M17 и М198. Уже общепризнанным является мнение, что данная гаплогруппа маркирует участие древних европеоидов в формировании генофонда населения Алтае-Саян (Derenko et al., 2006; Харьков, 2007) . Данная гаплогруппа была выявлена у древнего населения Таримского бассейна (ок.1 900 лет назад) (Li et al., 2010:9), у носителей пазырыкской культуры на Алтае (450 лет до н.э.) (Ricaut et al., 2004; Keyser et al., 20099) и у хунну Монголии (300 - 100 лет до н.э.) (Kim et al., 2010). Уже это показывает близость между древними популяциями данных регионов. Но, к сожалению, не было проведено генотипирование образцов на Y-STR, которое позволяет сравнивать разные образцы одной гаплогруппы. Образцы из захоронений андроновской, тагарской и таштыкской культур были определены Y-STR, что позволяет сравнить эти образцы между собой и образцами представителей разных популяций, как древних, так и современных.
Гаплотип андроновцев S10, S16 имеет следующий вид:
ANDRON S10, S16 13-25-16-11-11-14-10-14-11-18-15-14-11-16-20-12-23
При сравнении на 10 Y-STR гаплотипы андроновцев имеют полное совпадение с гаплотипами представителей следующих популяций: южные алтайцы (рода тодош, кергиль, тонжоан, чапты, мундус) (17), алтайцы (2), киргизы (65), тувинцы (4), монголы (1), телеуты (1), иранцы (1), индийцы (3), турки (1), русские (3), поляки (4), румыны (1), русины (1), словаки (2), англичане (1).
[Для сравнения использовались следующие базы данных, содержащих разномаркерные гаплотипы: База данных лаборатории эволюционной генетики НИИ медицинской генетики ТНЦ СО РАМН (TL), Family Tree DNA (FTDNA), Sorensen Molecular Genetic Foundation (SMGF), Y Chromosome Haplotype Reference Database (YHRD), и гаплотипы, приведенные в работах: Derenko et al, 2006; Roewer et al., 2008; Haber et al., 2010; Järve et al., 2009; Balanovsky et al., 2011]
Это всего 97 из 13 000 гаплотипов R1a1a, с которыми сравнивались гаплотипы андроновцев. Наибольшее число совпадений наблюдается с тяньшанскими киргизами и южными алтайцами. Полные совпадения гаплотипов у популяций, которые территориально близки и имеют общую историю, возможно только в случае их генетического родства, случайные совпадения маловероятны.
Таким образом, южные алтайцы и тяньшанские киргизы являются потомками близких родственников енисейских андроновцев, вероятнее всего, потомками алтайских андроновцев. Как известно, между киргизами и южными алтайцами лингвистами и этнографами давно было установлено близкое языковое и этническое родство (Баскаков,1966: 15-16). У этих народов совпадает большинство названий родовых подразделений (мундус, тёлёс – доолос, кипчак, найман, меркит и т.д.). Киргизские предания называют Алтай прародиной своего народа. Ряд историков считает, что некогда киргизы и южные алтайцы входили в единую общность и переселение киргизов с Алтая на Тянь-Шань произошло относительно недавно (Абрамзон, 1959: 34; Абдуманапов, 2007:95, 114). Модальный 15-маркерный гаплотип одной из групп южных алтайцев гаплогруппы R1a1a, приведенный в работе О.А.Балаганской (Балаганская, 2011: 22) , полностью совпадает с модальным гаплотипом самой многочисленного кластера R1a1a у тяньшанских киргизов.
13-25-16-11-10-11-11-18-15-8-11-16-21-12-23
[Порядок маркеров: DYS393 DYS390 DYS19 DYS391 DYS439 DYS389I DYS392 DYS389II DYS 458 DYS 437 DYS 438 DYS 456 DYS 448 DYSH4 DYS 635]
Особенно важным является значение маркера DYS448, равное 21. Это значение является главным отличительным признаком алтайско-киргизского кластера R1a1a, у большинства представителей этой гаплогруппы значение этого маркера равно 20. База SMGF содержит значительное количество киргизских гаплотипов R1a1. Количество гаплотипов со значением 21 и 22, намного превышает количество гаплотипов со значением 20. Все гаплотипы из этой базы, совпадающие на 12 маркерах с андроновскими, имеют значение DYS448 = 21, т.е. принадлежат к алтайско-киргизскому кластеру R1a1a.
При сравнении на 16 маркерах отличие на одну мутацию имеют гаплотипы тагарцев, исследованных в вышеуказанной работе Кейзер и обозначенных как S28 и S29, киргизы (5), иранцы (2), индиец (1), русские (1), словаки (2), венгры (1), поляки (1).
Это всего 15 из 8 тысяч гаплотипов. При этом словак (Dutzman), венгр (Meszaros) и поляк (Berezik) принадлежат к одному, т.н. южнорусскому кластеру R1a1a, киргизы к алтайско-киргизскому кластеру, индиец принадлежит к субкладу R1a1a, определяемый наличием SNP-маркёра L342.2, в состав которого входит и алтайско-киргизский кластер.
Совсем недавнее открытие нового SNP-маркёра L342.2 у гаплогруппы R1a1a позволяет в значительной мере прояснить ситуацию с происхождением и родственными связями представителей азиатских кластеров этой гаплогруппы. Обнаружен был этот SNP-маркёр и у представителей алтайско-киргизского кластера R1a1a1.
Предварительные результаты следующие: ареал распространения SNP-маркёра L342.2 оказался хоть и значительным, но все же не выходящим за рамки Азии. В Европе этот SNP-маркёр практически не встречается, за исключением населения явно азиатского происхождения: евреев-ашкенази и литовских и поволжских татар. Этот SNP-маркёр чаще встречается у следующих групп населения: арабы, проживающие в основном на пограничье с Ираком, турки, пакистанцы, северные и южные индийцы, афганцы, южные алтайцы, тяньшанские киргизы и башкиры. Близкими к протестированным представителям субклада R-L342.2 являются иранцы и иракцы, 44-маркерные гаплотипы которых представлены в базе SMGF. У казахов пока этот SNP-маркёр не обнаружен. Но часть казахов
1 [В настоящее время генотипирование на наличие L342.2 осуществляется в коммерческой лаборатории (Family Tree DNA), ни одна научная лаборатория еще не начала работу по выявлению этого SNP-маркёра. Это затрудняет возможность точно очертить ареал распространения этой субгаплогруппы, так как среди тестируемых в основном оказываются жители более развитых регионов.]
по количеству мутаций приближаются к основным кластерам субклада R-L342.2. Обнаружение этого SNP-маркёра у казахов позволит включить и Западный Казахстан в ареал этого субклада.
Основываясь на столь близком совпадении гаплотипов андроновцев и представителей алтайско-киргизского кластера, который относится к субкладу R-342.2, можно с большой долей уверенности предположить, что гаплотипы андроновцев также относятся к этому же субкладу.
Достаточно сложным является вопрос о прародине субклада R-L342.2. Однозначно можно сказать, что алтайско-киргизский кластер и, следовательно, саяно-алтайские андроновцы не являются предковой группой для всех остальных представителей этого субклада. Сравнение
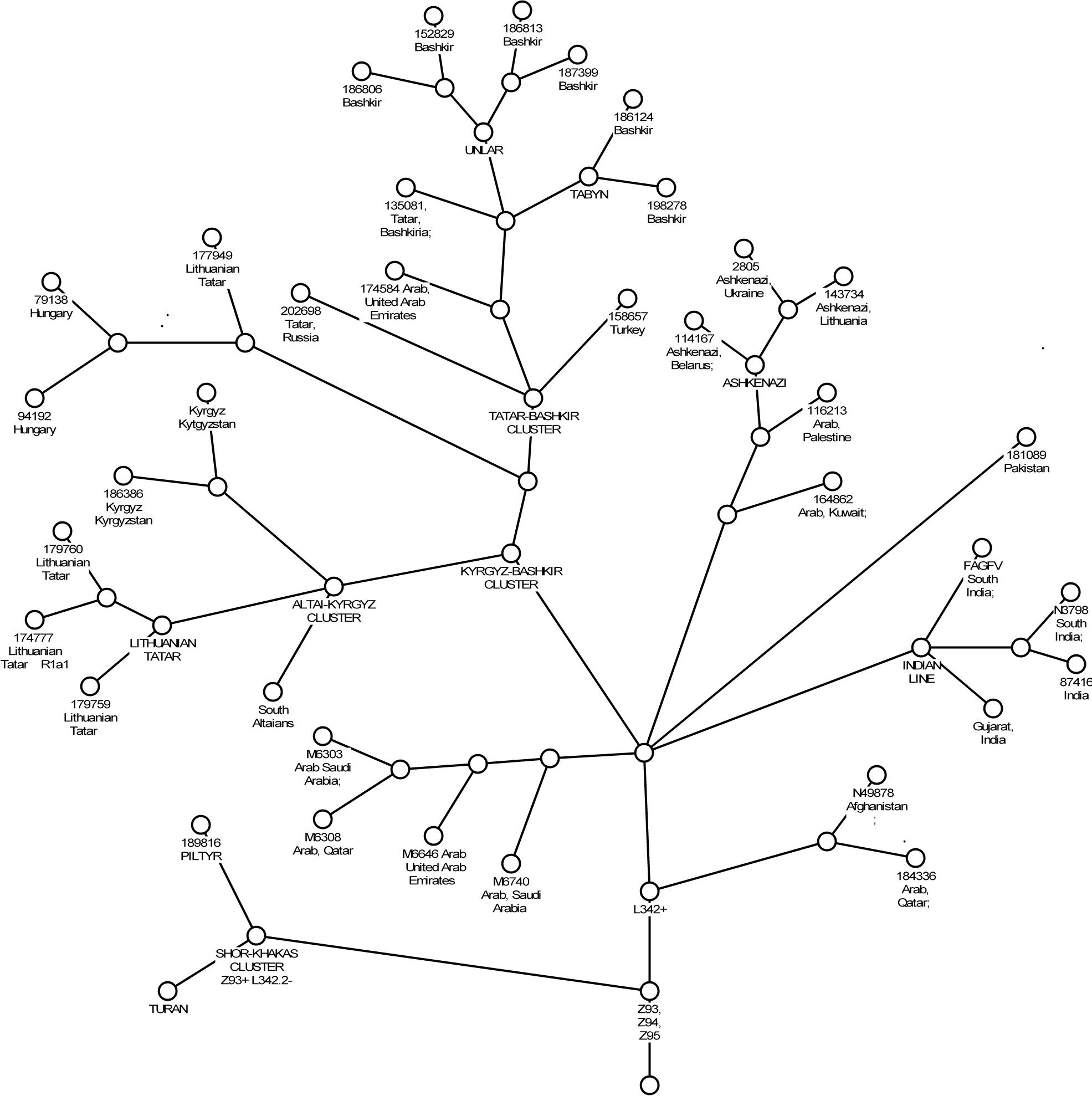
Ил.2. Филогенетическое древо субклада R-L342.2
гаплотипов показывает и значительно разнообразие внутри этого субклада и, следовательно, значительный возраст. Приблизительный возраст этого субклада, определенный нами, составляет 4500 – 5000 лет назад (3000 – 2500 лет до н.э.).
Филогенетическое древо (ил.2) позволяет определить наиболее вероятную прародину субклада R-L342.2. Это пограничье Ирана и Афганистана. Т.е. восточная часть Передней Азии. Возможно, в состав этой прародины входила и территория Южного Туркменистана.
Территория распространения субклада R-L342.2. в его южной части (Турция, Иран, Афганистан, Пакистан, Индия) слишком явно совпадает с современным ареалом распространения индо-иранских языков, так что этому нельзя не придать особого значения. С большой долей вероятности можно также предполагать, что распространение индо-иранских языков в этом регионе связано именно с субкладом R-L342.2. При этом, не вызывает практически никакого сомнения, что представители этого субклада были основой тех индо-ариев, которые «вторглись» в Индию примерно 3500 лет назад (Бонгард-Левин, Грантовский, 1983: 122, 123) . Значительная часть арабов этого субклада происходит из племени Тамим, которое пришло в Аравию из Ирака и до принятия ислама исповедовало зороастризм (Foltz, 2007: 95; Lecker. 2005: 71-75) , что свидетельствует о иранском происхождении этого племени.
Таким образом, наблюдается несомненная генетическая связь населения восточного ареала андроновской культуры (Саяно-Алтай, Семиречье) с территорией Ирака, Ирана, Афганистана и Индии. В этом аспекте наиболее вероятной представляется миграция мужских предков енисейских и алтайских андроновцев с юга Средней Азии. (Более поздние миграции потомков андроновцев в обратном направлении естественно происходили, об этом свидетельствует немало источников, но, видимо, они не оставили столь заметных генетических следов).
Близкие данные дают исследования одонтологических особенностей андроновцев Алтая. «Андроновцы Алтая в одонтологическом плане имеют явные черты южного происхождения. Одонтологический комплекс населения андроновской культуры Алтая не обнаруживает влияния со стороны монголоидного населения и проявляет сходство с европеоидными одонтотипами южного происхождения. По величине расстояний Снита палеопопуляции с территории Туркменистана (Гонур) и андроновской культуры Алтая сближаются с современными группами южных европеоидов. Суммируя результаты исследования одонтологического комплекса населения андроновской культуры Алтая, можно заключить, что он имеет южное происхождение. Результаты изучения одонтологических признаков, которые наследуются независимо от краниометрических, показывают, что население андроновской культуры Алтая было генетически связано с европеоидами южного происхождения» (Тур, 2009: 233-235).
Значимым является сближение с населением Гонур-депе. Бактрийско-Маргианская цивилизация, к которой относится Гонур-депе, занимает важное место в истории распространения индо-иранских языков. Согласно «Бактрийско-маргианской» гипотезе, ареал первоначального распространения индо-иранских языков находился в зоне бактрийско-маргианской культуры конца III – начала II тыс. до н. э. на юге Средней Азии и в Афганистане (Сарианиди, 1977, 1993) . Датировки культур хорошо соотносятся с возрастом субклада R-L342.2. Бактриано-Маргианский археологический комплекс (Гонур-депе) – III тысячелетие до н.э. (5000 – 4000 лет назад). Андроновские памятники Минусинской котловины датируются 1800–1400 гг. до н.э. Т.е. 3800 – 3400 лет назад.
Андроновскую культуру, как правило, относят к индоиранцам (Денисов, 2001: 4-21). . Е.А.Хелимский считает, что андроновцы не являются предками индийцев и иранцев, а в языковом отношении находятся примерно в одинаковой степени родства с нуристанцами, индийцами и иранцами, и считают представителей андроновской культуры носителями четвёртой, исчезнувшей ветви индоиранских языков. Он также придерживается версии о нахождении общей прародины индо-иранцев и андроновцев в районе Хорезма (Мерв, Герат). При явной близости андроновцев к индо-иранцам, андроновская культура моложе древних индоиранских (Миттани, Хорезм). По мнению У.А.Хелимского, эти данные, а также ряд других указывают на приход андроновцев с юга (Хелимский, 1998: 503 – 505). Все это в большой степени согласуется с генетическими данными.
Расчеты по количеству мутаций на 12-маркерных и 67-маркерных гаплотипах дают следующую картину. Наиболее близкими к субкладу R-L342.2 являются казахские гаплотипы и гаплотипы, принадлежащие к южнорусскому кластеру R1a1, в который попадают и несколько казахских и башкирских гаплотипов. Представители южнорусского кластера в основном проживают в Южной России (от Орловщины на севере до Северного Кавказа на юге), часть встречается в Польше. У представителей южнорусского кластера отсутствует SNP-маркёр L342.2, а казахов еще не тестировали на наличие этого SNP-маркёра.
Таким образом, вырисовывается достаточно обширный ареал, разные группы населения которого ныне и, вероятнее всего, в древности, были генетически близки. Большинство генетических мужских линий составляли представители гаплогруппы R1a1a. Но в этот ареал не входит Центральная Европа, Украина и Белоруссия, хотя представители этой гаплогруппы и в древности и ныне составляли и составляют большинство мужского населения этих регионов, но кластеры R1a1 на этой территории более удалены от субклада R-L342.2, чем южнорусский кластер.
Ямная, катакомбная и срубная культуры, располагавшиеся в Южной России, Поволжье и Приуралье, имеет гораздо больше сходства с андроновской в ее алакульском варианте, чем с находящейся западнее культурой шнуровой керамики и т.п. Пожалуй, этот факт не оспаривается ни археологами, ни антропологами (Гинзбург, 1959, 1962; Трофимова, 1961; Алексеев, 1964, 1967). Ямную культуру даже считают предковой и для срубной и для андроновской. Недаром выделяется территория скифо-сибирского мира, сложившийся на основе ямной, катакомбной, срубной и андроновской культур (Мартынов, 2000)
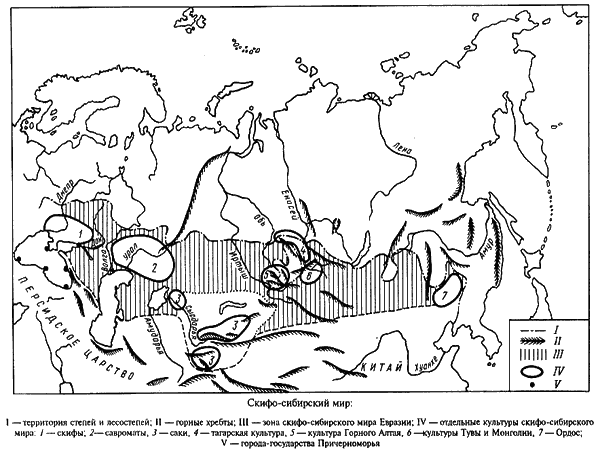
Ил.3. Скифо-сибирский мир. (Мартынов, 2000)
Доминирование грациального одонтологического типа наблюдается у восточных ямников. А.В.Зубова указывает, что истоки морфологических особенностей этого грацильного субстрата лежат в среде населения Малой, Передней и Средней Азии. Представленные в ее работе результаты кластеризации частот одонтологических признаков носителей разных археологических культур убедительно показывают, что наибольшее сближение даже у украинских ямников наблюдается с культурами Туркмении (Алтын-депе, Сапалли-тепе) и Малой Азии (Чатал-Гуйюк) (Зубова, 2010. 93) .
По мнению большинства исследователей арийской проблематики, полукочевые скотоводческие племена срубной и андроновской культурно-исторических общностей представляют индоиранскую группу индоевропейской языковой семьи и возможно являются легендарными ариями, которые в середине II тыс. до н. е. проникли в древний Иран, перешли хребты Гиндукуша, горы Афганистана и вторглись в долину Инда. Абашевские племена также считаются представителями индоиранской языковой общности (Кузьмина, 1994, 2008.. Кузьмина, 1995. Бонгард-Левин, Грантовский, 1974) . По мнению А.Х.Халикова, многочисленные индоиранские лексические заимствования, которые выявлены у финно-угорских народов Евразии, особенно у восточно-волжских, пермских и угорских, возникли именно благодаря контактам с индо-иранских абашевской, срубной и андроновской культур с племенами лесостепи Восточной Европы и Западной Сибири (Халиков, 1990: 53).
Но значительной проблемой является выяснение направления древних миграций. Сторонники западноевропейской, а также южнороссийской прародины индоевропейских языков считают андроновскую культуру несомненно принадлежащий к кругу индоевропейских культур и, следовательно, придерживаются версии прихода предков андроновцев с запада из южнорусских степей. Этой версии придерживается и часть антропологов (Козинцев, 2008, 2010). . Генетические данные, рассмотренные выше, показывают скорее обратную картину, движение предков андроновцев с территории близкой к Юго-Западной Туркмении. На данном этапе исследования еще непонятно, откуда пришли предки андроновцев и индо-иранцев кластера R-L342.2 на их общую прародину. Не исключены и северный и южный маршруты.
Возможно, точку в этом вопросе поставит нахождение прародины всей гаплогруппы R1a1a. Часть исследователей придерживается мнения о нахождении таковой на Балканах, часть в Южной России, часть помещает ее Передней Азии или даже в Северо-Западной Индии. К сожалению, в настоящее время эта проблема еще не решена, соответственно не решена проблема первичной прародины индо-иранцев и истоков андроновской культуры, но генетика, в отличие от археологии, антропологии и лингвистики обладает гораздо большими возможностями для решения этого вопроса. Выявление новых SNP-маркёров и анализ древних ДНК в ближайшее время позволит ответить на большинство подобных вопросов.
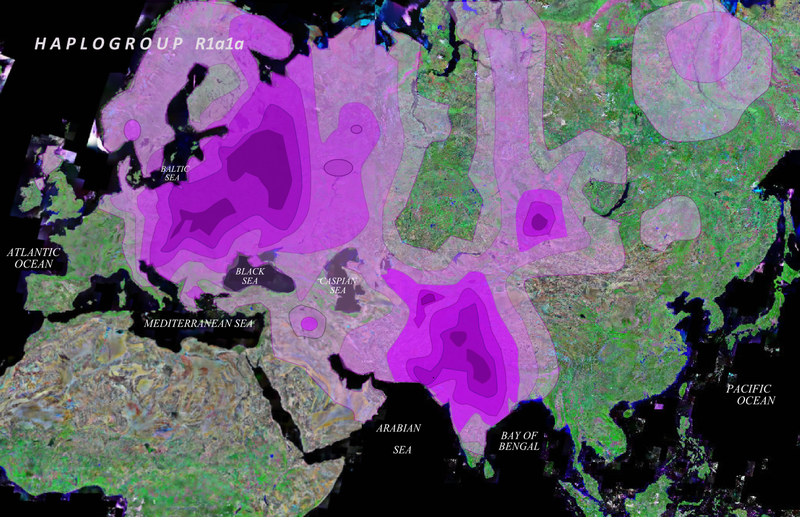
Ил.4. Распространение гаплогруппы R1a1a (по Underhill P. et al.:). Темнофиолетовым цветом выделены повышенные частоты этой гаплогруппы.
Возможны два варианта. Миграция предков субклада R-L342.2 из Восточной Европы или миграция из Западной Азии, района присутствия предковых для R1a1 гаплогрупп, но зарождение этого субклада все равно должно быть связано с пограничьем Ирана, Афганистана и Средней Азии. Также можем предполагать, что распространение индо-иранских языков, в том числе на территорию Восточной Европы шло из этого или близкого региона.
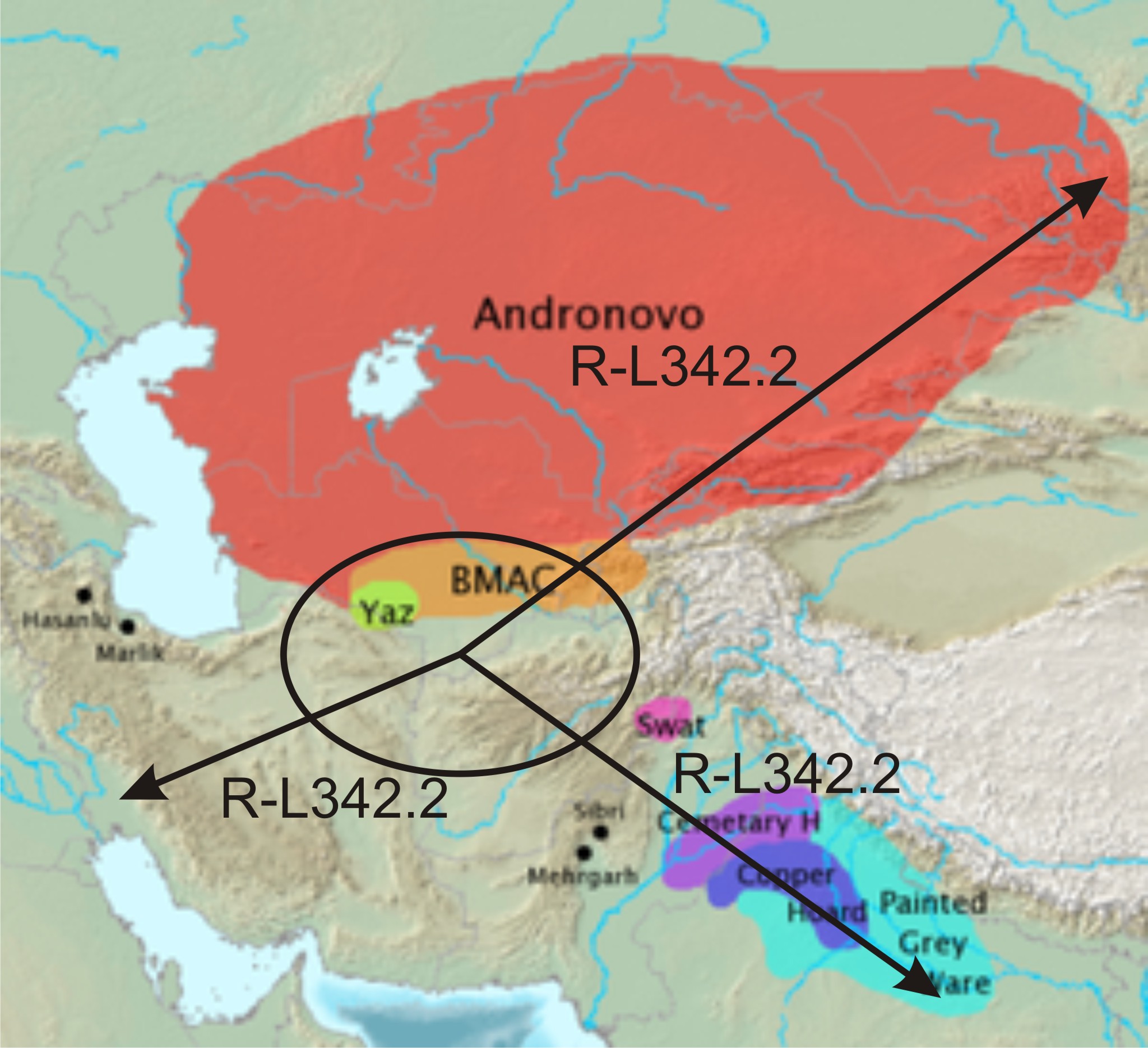
Ил.5. Миграции субклада R-L342.2
BMAC – ареал Бактрийско-Маргианской археологической культуры.
В вышеуказанной работе Кайзер также представлены гаплогруппы и гаплотипы носителей тагарской и таштыкской культур, погребенных в следующих захоронениях:
№7. S24 – Хакассия – Тагарская культура - 800 г. до н.э. –100 г. н.э.
№9. S25 – Хакассия - Тагарская культура - 800 г. до н.э. –100 г.
№10. S26 – Бейский район, Хакассия - Тагарская культура - 800 г. до н.э. –100 г.
№11. S28 и S29 – Богратский район, Хакассия - Тагарская культура - 800 г. до н.э. –100 г.
№5 - S32 и S34 – Богратский район, Хакассия - Тагарская культура - 800 г. до н.э. –100 г.
Гаплотип представителя тагарской культуры, обозначенного как S28, принадлежит также к гаплогруппе R1a1a и имеет следующий вид:
TAGAR S28 13-25-16-11-11-14-10-14-11-17-15-14-11-16-20-12-23
Гаплотип этого тагарца ближе к предковому гаплотипу всех R1a1a и, поэтому, имеет больше совпадений среди гаплотипов, представленных в разных базах данных. Среди встреченных близких совпадений: алтайцы южные (чапты, тонжоан, мундус) (3), телеуты (из рода тодош) (3), киргизы (8), монголы (2), венгры (5), ливанцы (1) и значительное число представителей южнорусского кластера R1a1a. Какие-либо выводы о происхождении предков носителей этих гаплотипов сделать сложно из-за частой встречаемости.
На 16 маркерах наблюдается полное совпадение только с гаплотипами венгра, поляка и словака. Все три гаплотипа относятся к южнорусскому кластеру.
Не исключено, что это указывает на связь с территорией южнорусских степей, при этом эти венгры и словаки могут быть потомками мадьяр – относительно недавних переселенцев на запад и являться представителями восточной ветви южнорусского кластера, а сам южнорусский кластер – потомками населения срубной культуры.
Другой представитель тагарской культуры имеет следующий гаплотип:
TAGAR S29 13-25-x-11-11-14-x-14-11-17-15-14-11-16-x-12-23
Этот гаплотип имеет недостаточное для анализа количество маркеров, но его 14-маркерный формат идентичен формату предыдущего.
Другие гаплотипы представителей тагарской и таштыкской культур оказались очень редкими.
TAGAR S26 13-24-16-11-11-14-10-13-11-18-15-14-11-16-20-13-23
Полные совпадения на 10 маркерах у данного образца наблюдаются с представителями следующих популяций: тувинцы (10), киргизы (5), хакасы (2), сойоты (1), тоджинцы (1), индийцы (1), турки (1), иранцы (1), украинцы (1), поляки (1), словаки (1), шведы (1).
Совпадений не так уж много. Всего 26 из 13 тысяч гаплотипов.
Следующий гаплотип TAGAR S25 13-24-x-11-11-14 -10-13-11-18-15-14-11-16-20-x-23 Этот гаплотип также имеет недостаточное для анализа количество маркеров, но его 15-маркерный формат идентичен формату предыдущего.
Гаплотипы представителей тагарской культуры (TAGAR S24) и таштыкской (TASTYK S34) полностью идентичны. Отличаются от тагарца S26 только на одну мутацию 13-24-17-11-11-14-10-13-11-18-15-8-11-16-20-13-23
Гаплотип S24 и S34 является производным от гаплотипа тагарца S26. Полное совпадение только с гаплотипом одного киргиза. Близкими оказываются гаплотипы одного индийца и туркмена.
Несомненно, что таштыкец S34 – близкий родственник тагарца S24. Появление таштыкской культуры связывают с миграцией в Минусинскую котловину носителей другого антропологического облика. Таким образом, данный представитель таштыкской культуры не является потомком мигрантов, как следовало ожидать, а скорее одним из прямых потомков тагарцев, которые, несмотря на смену культуры, продолжали существовать. При этом не исключено, что исследования останков других носителей таштыкской культуры выявило бы другие гаплогруппы.
Гаплотип еще одного представителя тагарской культуры имеет следующий вид TAGAR S32 13-24-17-11-11-14-10-13-12-18-15-8-11-16-20-13-23
Данный гаплотип является явно производным от предыдущего (S24, S34), полных совпадений нет.
Совпадения гаплотипов киргизов, принадлежащих к разным кластерам, не только с гаплотипами андроновцев, но и тагарцев и таштыкца однозначно
указывает на наличие общей предковой популяции. При этом среди киргизских гаплотипов выявлено лишь незначительное число общих совпадений с гаплотипами современных хакасов. Это доказывает, что миграции енисейских киргизов на территорию Киргизстана хотя и имели место, но были редки и не повлияли значительно на генофонд тяньшанских киргизов. Как ни странно, но мы не находим явных потомков енисейских андроновцев и тагарцев среди современного населения Хакассии. Чаще наблюдается сближение с тувинцами, и еще в большей степени тяньшанскими киргизами и южными алтайцами. Требуются дальнейшие исследования, чтобы объяснить эту ситуацию.
Гаплотипы тагарцев при сравнении с гаплотипами андроновцев показывают следующее. Тагарские гаплотипы S28 и S29 часто встречаются в разных популяциях и могут быть как предковыми для андроновских, так и производными от них.
Гаплотипы андроновцев S10, S16 и тагарцев S24, S25, S26, S34, S32 являются производными от гаплотипа 13-25-16-11-11-14-10-13-11-18-15-14-11-16-20-12-23 который является предковым как для гаплотипов андроновцев, так и для гаплотипов тагарцев и таштыкца.
Можно представить родственные отношения носителей этих гаплотипов таким образом.
Ил.6.
В данном случае это показывает, что тагарцы не являются прямыми потомками енисейских андроновцев, а скорее указывает на то, что происхождение этих двух культур связано с одной предковой популяцией, располагавшейся юго-западнее.
Наблюдается сходство тагарских гаплотипов с гаплотипами представителей нескольких племенных групп башкирского этноса – табын и унлар. Легендарные предания этих племен выводят их с Алтая, точнее с пограничья Алтая, Тувы и Монголии (Кузеев, 1974: 254). И еще большее сходство наблюдается с другим, очень близким к башкирскому кластером внутри субклада R-L342.2, который представлен венграми из специфической этнической группы Шекели и татарами (литовским и поволжским). (см.ил.2). Модальный гаплотип этого кластера 13-24-17-11-11-14-12-12-11-14-11-18 отличается от тагарского S24 и таштыкского S34 на одну мутацию. У южных алтайцев также встречаются близкие гаплотипы 13-24-17-10-11-13-12-12-10-12-11-18
Наибольшее сближение татаро-венгерского и башкирского кластеров субклада R-L342.2 наблюдается с алтайско-киргизским кластером и гаплотипами поволжского татарина и турка (см.ил.2). К алтайско-киргизскому кластеру принадлежат в основном представители правого и левого крыла киргизских племен, которые считаются потомками Долон-бия, и которые, согласно родословным, являются выходцами с Алтая. Таким образом, в субкладе R-L342.2 выделяется северная ветвь, которую можно связать с андроновцами и тагарцами и уточнить точку расхождения тагарской ветви с основной андроновской. Совокупность разных данных указывает на территорию Горного Алтая.
Древние популяции, даже при явном доминировании одной гаплогруппы, вряд ли были полностью моногаплогруппными. Они могли включать представителей разных мужских гаплогрупп, которые, естественно, мигрировали совместно. Среди киргизов и алтайцев встречаются представители гаплогрупп, происхождение которых из Передней Азии не вызывает сомнения. Переднеазиатские гаплогруппы J, E1b1b1 и G с небольшими частотами встречаются по всей территории Алтая, не превышая 2%, за исключением гаплогруппы G, которая обнаруживает новый максимум на территории Средней Азии у некоторых групп казахов (Балаганская, 2011: 17.) . Возможно, миграции представителей этих гаплогрупп на Алтай происходили также в андроновскую эпоху.
Наблюдается постоянные параллели в распределении субклада R-L432.2 и гаплогруппы R1b1b1(M73). Она часто встречается у алтайцев (телеуты, кумандинцы), шорцев, тувинцев, казахов (род кыпшак), восточных башкир, также у поволжских татар, киргизов и уйгур (FTDNA; SMGF; Балаганская, 2001: 10; Myres, 2010: 97). . Первичные миграции этой гаплогруппы связаны с
югом Средней Азии. Гаплогруппа Q1a3 также зародилась в регионе, близком к Ирану и Афганистану. Она является предковой для гаплогруппы Q1a3a, распространенной среди американских индейцев. Видимо, эта гаплогруппа была одной из первых мужских гаплогрупп, появившихся в Сибири. Встречается со значительной частотой у кетов, селькупов, тувинцев, алтайцев, а также у киргизов. Один из субкладов этой гаплогруппы встречается в Скандинавии (TL, FTDNA, SMGF).
Таким образом, значительное число миграций мужских гаплогрупп, из которых Q1a3 и R1b1a1 уже в глубокой древности присутствовали в Южной Сибири, шли в направлении Иран, Афганистан –> Средняя Азия –> Южная Сибирь. Без сомнения, представители всех этих гаплогрупп участвовали в формировании генофонда андроновцев и тагарцев. Нужно учесть, что на антропологический облик, а также на генофонд и культуру андроновцев и тагарцев могли повлиять и представители мужских гаплогруппы восточноевразийского происхождения C и N, одна из которых даже встретилась у андроновца.
Следует оговориться, что выше мы рассматривали исключительно миграции мужских генетических линий. На генофонд андроновцев и тагарцев несомненное влияние оказали и женские гаплогруппы, часть из которых присутствовала в Южной Сибири с древнейших времен, часть мигрировала вместе с мужскими предками андроновцев. Женские гаплогруппы, выявленные у тагарцев и таштыца (U5a1, U4a, I4, C, H5a, HV и т.д.) показывают включение в состав тагарцев женских линий разного происхождения – и восточноевразийских и западноевразийских. В составе генофонда современных тюрков Южной Сибири достаточно многого женских гаплогрупп и специфических субкладов, происхождение которых связано территорией Западной Азии и Индийского субконтинента (Деренко, 2009: 26). .
Все это создает достаточно сложную картину формирования и генофонда и культуры андроновцев и тагарцев.
Выводы.
Гаплотипы носителей андроновской и тагарской культур имеют наибольшее близость к гаплотипам южных алтайцев и тяньшанских киргизов.
Наблюдается близкая генетическая связь между представителями гаплогруппы R1a1a, населяющими территорию Средней, Центральной, Южной и Передней Азии. Эта связь обусловлена принадлежностью к одному субкладу R-L342.2.
Cубклад R-L342.2 является общим компонентом, присутствующим у современных тюркских и индо-иранских народов.
Основываясь на генетических данных, можно утверждать, что носители андроновской и тагарской культур являются представителями субклада R-L342.2.
Генетические данные подтверждают данные других наук, что андроновская и тагарская культура имеет близкую генетическую связь с предками индо-иранских народов.
Наиболее вероятно происхождение носителей андроновской и тагарской культур с территории пограничья Ирана, Афганистана и Средней Азии.
К сожалению, в настоящее время результаты исследования древней ДНК еще редки. Также еще не выявлены SNP-маркёры, позволяющие с большей точностью проанализировать расположение разных субкладов гаплогруппы R1a1 и, таким образом, выяснить направление древних миграций ее представителей.
Выявление наличия SNP-маркёра L342.2 у носителей андроновской и тагарской культур могло бы полностью подтвердить наши предположения о южном происхождении андроновцев и тагарцев, показать степень родства с индо-ариями и другими древними индо-иранскими народами, а также найти подлинных генетических потомков носителей этих культур.
Массовое генотипирование на этот SNP-маркёр в научных лабораториях представителей разных популяций, как современных, так и древних, позволит ответить на многие вопросы древней истории Южной Сибири и Средней и Центральной Азии.
Волков Владимир Геннадьевич
Зав. Экспозиционным отделом МУ «Музей истории Томска»
Харьков Владимир Николаевич
Кандидат биологических наук, старший научный сотрудник лаборатории эволюционной генетики НИИ Медицинской генетики ТНЦ СО РАМН
Степанов Вадим Анатольевич
Доктор биологических наук, профессор, заместитель директора по научной работе, руководитель лаборатории эволюционной генетики НИИ Медицинской генетики ТНЦ СО РАМН
162
__________________________________________________________
Базы данных Y-STR гаплотипов:
Family Tree DNA (FTDNA) Электронный ресурс. Доступ: [url]http://www.familytreedna.com[/url],
Sorensen Molecular Genetic Foundation (SMGF) Электронный ресурс. Доступ: [url]http://www.smgf.org[/url]
Y Chromosome Haplotype Reference Database (YHRD) Электронный ресурс. Доступ: [url]http://www.yhrd.org[/url]
Литература
Balanovsky O., Dibirova K., Dybo A. et al. Parallel Evolution of Genes and Languages in the Caucasus Region // Molecular Biology and Evolution, 2011: 1 – 49. Приложения: Электронный ресурс. Доступ:[url]http://mbe.oxfordjournals.org/content/early/2011/05/13/molbev.msr126/suppl/DC1[/url]
Chikisheva T.A., Gubina M.A., Romaschenko A.G., Karafet T.M. et al.//Archaeology, Ethnology and Anthropology of Eurasia, 2007, 4 (32): 130-142.
Derenko M., Malyarchuk B., Denisova G. et al. «Contrasting patterns of Y-chromosome variation in South Siberian populations from Baikal and Altai-Sayan regions»//Human Genetics, 2006. 118: 591-604.
Gao S, Cui Y, Yang Y et al.Mitochondrial DNA analysis of human remains from the Yuansha site in Xinjiang, China.// Sci China C Life Sci., 2008 Mar. 51(3):205-13.
Foltz R. L'Iran, creuset de religions: de la préhistoire à la République islamique.- Laval, 2007.- 170 p.
Haber M., Platt D.E., Badro D.A. et al. Influences of history, geography, and religion on genetic structure: the Maronites in Lebanon.//European Journal of Human Genetics, 2011, 114 (2): 334-40.
Järve M., L.A. Zhivotovsky, S. Rootsi et al. Decreased Rate of Evolution in Y Chromosome STR Loci of Increased Size of the Repeat Unit.// PLoS ONE 2009; 4: e7276 (open access).
Приложения: Электронный ресурс. Доступ:[url]http://www.ncbi.nlm.nih.gov/pmc/articles/PMC2748704/bin/pone.0007276.s001.xls[/url]
Kim K., Brenner C.H., Mair V.H., et al. A Western Eurasian Male Is Found in 2000-Year-Old Elite Xiongnu Cemetery in Northeast Mongolia.//American Journal of Physical Anthropology, vol. 142, no. 3 (July 2010), pp. 429–440.
Keyser C. et all. Ancient DNA provides new insights into the history of south Siberian Kurgan people.//Human Genetics, 2009. Vol.126, Num. 3, 395-410
Keyser-Tracqui C., Crubezy E., Ludes B. Nuclear and Mitochondrial DNA Analysis of a 2,000-Year-Old Necropolis in the Egyin Gol Valley of Mongolia.// Am.J.Hum.Genet, 2003, 73: 247-260.
Krause J, Briggs AW, Kircher M, Maricic T, Zwyns N, et al. Complete mtDNA genome of an early modern human from Kostenki, Russia.//Current Biology, 2009, Volume 20, Issue 3, 231-236.
Lecker, Michael. People, tribes, and society in Arabia around the time of Muḥammad. - Burlington.- 2005.- 350 p.
Li C., Li H., Cui Y. et al. Evidence that a West-East admixed population lived in the Tarim Basin as early as the early Bronze Age.//BMC Biology 2010, 8:15:1-12.
Malyarchuk B., Derenko M., Denisova G., et al. Phylogeography of the Y-chromosome haplogroup C in northern Eurasia // Annals of Human Genetics. 2010. 74(6): 539-46.
Приложения: Электронный ресурс. Доступ: [url]http://malyarchuk-bor.narod.ru/Table_S1.xls[/url]
Myres et al. A Major Y-chromosome Haplogroup R1b Holocene Era Founder Effect in Central and Western Europe // European Journal of Human Genetics (2011) 19, 95–101.
Ricaut F.X., Keyser-Tracqui C., Cammaert L. et al. Genetic analysis and ethnic affinities from two Scytho-Siberian skeletons.//American Journal of Physical Anthropology, vol. 123, no. 4 (Apr 2004), pp. 351-60.
Roewer L. et all. (2008). Analysis of Y chromosome STR haplotypes in the European part of Russia reveals high diversities but non-significant genetic distances between populations. Int. J. Legal Medicine 122 (3), 219 – 223.
Приложения: Электронный ресурс. Доступ: [url]http://www.ncbi.nlm.nih.gov/pmc/articles/PMC2755792/bin/414_2007_222_MOESM1_ESM.xls[/url]
Schilz F. Molekulargenetische Verwandtschaftsanalysen am prähistorischen Skelettkollektiv der Lichtensteinhöhle, Dissertation, Göttingen 2006, 252 s.
Underhill P., Myres N.M., Rootsi S. et all. Separating the post-Glacial coancestry of European and Asian Y chromosomes within haplogroup R1a // European Journal of Human Genetics, 2009. 18(4): 479.
Zhao YB, Li HJ, Cai DW et al. Ancient DNA from nomads in 2500-year-old archeological sites of Pengyang, China.// Journal of Human Genetics 55, April 2010, 215-218.
Абдуманапов Р.А. Кипчакский компонент в этногенезе киргизов. Диссертация на соискание ученой степени кандидата исторических наук.- Томск – 2007: 183 с.
Абрамзон С.М. Вопросы этногенеза киргизов по данным этнографии // Труды Киргизской археолого-этнографической экспедиции. – Фрунзе: Издательство Академии наук Киргизской ССР, 1959. – Т. 3. – С. 31-44.
Алексеев В.П. Антропологический тип населения западных районов распространения андроновской культуры.- В кн.: Проблемы этнической антропологии Средней Азии.- Ташкент, 1964.- С.19 – 28.
Алексеев В.П. Антропология андроновской культуры.- Советская Археология.- М.,1967.- №1.- С.22-26.
Балаганская О.А. Полиморфизм y хромосомы у тюркоязычного населения Алтая, Саян, Тянь-Шаня и Памира в контексте взаимодействия генофондов Западной и Восточной Евразии. Автореферат диссертации на соискание ученой степени кандидата биологических наук.- Москва, 2011.- 26 с.
Баскаков Н.А. Тюркские языки (общие сведения и типологическая характеристика) // Языки народов СССР. – М.: Наука, 1966. – Т. 2. – С. 15-16.
Бонгард-Левин Г. М., Грантовский Э. А. От Скифии до Индии. Древние арии: мифы и история.— М.: Мысль, 1974. — 206 с.
Волков В.Г., Харьков В.Н., Штыгашева О.В., Степанов В.А. Генетическое исследование хакасских и телеутских сеоков. Сравнительная характеристика по данным маркеров Y-хромосомы.// Культура как система в историческом контекте: опыт западно-сибирских археолого-этнографических совещаний.- Томск, 2010.- С.403 – 405.
Гинзбург В.В.Материалы к антропологии населения Западного Казахстана в эпоху бронзы.//Материалы и исследования по археологии СССР.- М.Л.,1962.- №120.- С.186 – 198;
Денисов И. В. Некоторые проблемы археологии бронзового века Волго-Уралья и Ведийско-Авестийские сказания // В центре Евразии: Сборник научных трудов. — Стерлитамак, 2001.- С. 4-21.
Деренко М.В. Молекулярная филогеография коренного населения Северной Азии по данным об изменчивости митохондриальной ДНК. Автореферат диссертации на соискание ученой степени доктора биологических наук.- М., 2009.- 49 c.
Зубова А.В. Население ямной культурно-исторической общности в свете одонтологических данных.// Вестник археологии, антропологии и этнографии. 2010. № 2 (13).- С.85 -95.
Козинцев, А.Г. Так называемые средиземноморцы Южной Сибири и Казахстана, индоевропейские миграции и происхождение скифов.//Археология, этнография и антропология Евразии.- 2008.- №6.- С.140 – 144.
Козинцев А.Г. Ранние индоевропейцы Сибири и Центральной Азии по данным антропологии.//Культура как система в историческом контекте: опыт западно-сибирских археолого-этнографических совещаний.- Томск, 2010.- С.419 – 421.
Кузеев Р. Г. Происхождение башкирского народа. Этнический состав, история расселения. М.: Наука, 1974. 570 с.
Кузьмина Е. Е. Арии — путь на юг. — М.: Летний сад, 2008. — 560 с.:
Кузьмина, Е. Е. Откуда пришли индоарии? : материальная культура племен андроновской общности и происхождение индоиранцев.- М., 1994. — 464 с.
Кузьмина О. В. Соотношение абашевской и покровской культур // Конвергенция и дивергенция в развитии культур эпохи энеолита-бронзы Средней и Восточной Европы. — СПб, 1995. — Ч. 2. — С. 27-51.
Мартынов А.И. Археология: Учебник для студентов, обучающихся по специальности «История». Изд. 4. – Москва: Высшая школа, 2000. – 439 с.
Сарианиди В.И. Древние земледельцы Афганистана.- М.,1977.- 172 с.
Сарианиди В.И. В поисках страны Маргуш.- М., 1993.- 362 с.
Трофимова Т.А. Черепа из могильника тазабагъябской культуры Кокча III.- В кн.: Материалы Хорезмской экспедиции.- М., 1961.- вып.5.- С.97 – 146.;
Тур, С. С. Одонтологическая характеристика населения андроновской культуры Алтая .// Известия Алтайского государственного университета. Серия: История. Политология. - 2009. - № 4/2. - C. 228-236.
Халиков А.Х. основные этапы урало-индоевропейских связей в Среднем Поволжье и Приуралье // URALO-INDOGERMANICA.- Ч.2, М., 1990.- С.53 – 59.
Харьков В.Н., Степанов В.А., Медведева О.Ф. и др. Различия структуры генофондов северных и южных алтайцев по гаплогруппам Y-хромосомы // Генетика. - 2007. - Т. 43. - № 5. - С. 675–687.
Хелимский Е.А. Южные соседи финно-угров: иранцы или исчезнувшая ветвь Ариев (арии-андроновцы)? - К 70-летию В.Н. Топорова,- М., 1998.- С.503 – 505.
____________________________________________________