В первой части очерка о коллизии популяционной генетики и ДНК-генеалогии профессор А.А. Клёсов рассказал о том, в чём состоят существенные различия между этими двумя областями научного знания. В частности, ДНК-генеалогия и популяционная генетика используют различную методологию, разные термины и принципиально разный расчётный аппарат, что приводит к кардинально различающимся выводам и историческим интерпретациям. Были приведены показательные примеры. Автор отметил и то, что сторонники попгенетики не брезгуют откровенно вненаучными приёмами дискредитации, приводя в пример печально известных Балановских и их некоторых единомышленников. Сегодня – продолжение очерка, поэтому слово Анатолию Алексеевичу…

Профессор А.А. Клёсов после удачной охоты
Продолжим сравнительное рассмотрение ДНК-генеалогии и популяционной генетики, их расчетные методы и характер выводов. Рассмотрим пример, имеющий важное историческое значение. Он имеет прямое отношение к легендарным ариям, которые по историческим сведениям (скорее, предположениям) перешли в Индию примерно 3500 лет назад. Это впервые было высказано, видимо, немецко-английским ученым-ориенталистом Максом Мюллером (1823-1900), и эта дата приводится в книге Гордона Чайлдса «Арии. Основатели европейской цивилизации» (1926). Он пишет: «…Ригведа была создана вскоре после 1400 года до н.э., и вторжение ариев в Индию относится примерно к тому же времени» (то есть примерно 3400 лет назад). Но в следующем абзаце Чайлдс продолжает – «Недавно вызов этой традиционной точке зрения был брошен сразу с двух сторон. Паргитер считает, что проникновение ариев в Индию началось задолго до составления ведических гимнов… Изучение генеалогий правителей привело упомянутого автора к заключению, что арии… вторглись в Индию скорее ближе к началу, чем к концу 2-го тысячелетия до н.э. …Однако в последние годы она была оспорена и с другой стороны. Брюннхофер и некоторые другие исследователи утверждают, что …проникновение ариев в Индию следует относить к гораздо более позднему времени, чем это обычно предполагается».
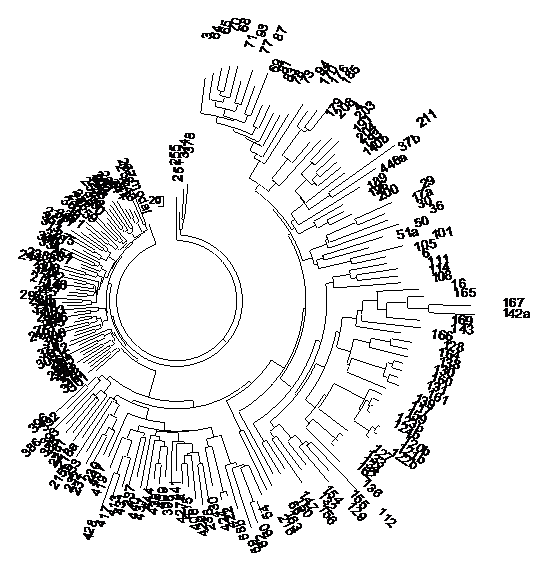
Рис. 1. Дерево 187 индийских гаплотипов Y-хромосомы в 37-маркерном формате. Слева – плотная ветвь из 64 гаплотипов гаплогруппы/субклада R1a-L657, с общим предком, жившим 4750±500 лет назад; справа и внизу – серии древних индийских гаплотипов гаплогрупп (по часовой стрелке) Н, L, Q, L, J-M304, O, C, L, C, J1-M267, J2b-L282, J2-M172, R2-M124, Q (повторы гаплогрупп – это отдельные ветви этих гаплогрупп). Общие предки ветвей справа и внизу жили 8-12 тысяч лет назад.
Построено по данным Индийского проекта FTDNA.
Слева – все 64 гаплотипа гаплогруппы R1a, которые образовали плотную ветвь, составленную из субклада R1a-L657. То, что ветвь вполне однородная, показывает следующее дерево, уже только из гаплотипов данной ветви R1a-L657. Однородность дерева уже показывает, что все гаплотипы родственные, все происходят от одного общего предка, во всяком случае, с той точностью, которая нас в данном случае вполне устраивает. Посчитаем, когда этот предок жил. Во всех 64 гаплотипах в 37-маркерном формате – 924 мутации, что дает 924/64/0.09 = 160 → 190 условных поколений, то есть 4750±500 лет назад. Расчет с помощью калькулятора Килина-Клёсова, основанного совершенно на другом принципе расчета, а именно по каждому маркеру отдельно, используя калиброванные абсолютные константы скорости мутаций для каждого из 37 маркеров, дал время жизни общего предка гаплотипов R1a в Индии 4964±548 лет назад, то есть, округляя, получаем 5000±550 лет назад. Это та же величина, что и полученная «линейным методом» 4750±500 лет назад, в пределах погрешности расчетов.
Рис. 2. Дерево 64 индийских гаплотипов гаплогруппы R1a-L657 в 37-маркерном формате. Общий предок дерева жил 4750±500 лет назад. Построено по данным Индийского проекта FTDNA.
Но это время, отдаляющее нас от времени жизни общего предка гаплотипов группы R1a, живущих в настоящее время в Индии. Это вовсе не значит, что этот предок жил в Индии 4750-5000 лет назад. В Индию его потомки пришли примерно 3400-4000 лет назад, то есть на тысячу лет позже времени жизни своего общего предка. Где же этот предок жил, откуда в Индию пришли арии?
Чтобы ответить на этот вопрос, рассмотрим базовый гаплотип индийских R1a. Он – следующий (к нему, ко временам примерно 5000 лет назад, сходятся все 64 указанные гаплотипа):
25 16 10 11 14 12 12 10 13 11 17 – 16 9 10 11 11 24 14 20 32 12 15 15 16 – 11 12 19 23 15 16 18 19 35 39 13 11
А вот – предковый гаплотип этнических русских гаплогруппы R1a, общий предок которых жил на Русской равнине 4900 лет назад. Гаплотип приведен в книгах «Происхождение славян» (А.А. Клёсов, 2013, стр. 26) и «Арийские народы на просторах Евразии» (А.А. Клёсов, К.А. Пензев, 2014, стр. 246):
25 16 11 11 14 12 12 10 13 11 17 – 15 9 10 11 11 24 14 20 32 12 15 15 16 – 11 12 19 23 16 16 18 19 35 38 14 11
Мы видим, что предковые гаплотипы ариев и этнических русских очень похожи, между ними на вид всего 5 мутаций. На самом деле там всего 3.495 мутаций, потому что все отличающиеся аллели дробные. 3.495 мутаций между двумя 37-маркерными базовыми гаплотипами разделяют эти гаплотипы (то есть предков индийцев и этнических русских) на 3.495/0.09 = 39 → 41 условных поколений, или примерно 1025 лет. То есть их общий предок (субклад R1a-Z645) жил примерно за 500 лет до появления общих предков сегодняшних этнических русских гаплогруппы R1a и индийцев той же гаплогруппы, или примерно 5500 лет назад.

Примерно ту же датировку дают снипы (то есть SNP, необратимые мутации в Y-хромосоме). По данным Владимира Таганкина, который серьезно занимается снипами Y-хромосомы, между Z645 и Z283 всего два снипа, между Z283 и Z282 всего один снип, и между Z282 и Z280 – всего три снипа. «Всего» – потому что по предварительным данным один снип в среднем образуется каждые 100-150 лет назад, данные еще недостаточно откалиброваны. Поэтому при шести снипах между Z645 и Z280 (последний образовался примерно 5000 лет назад), Z645 (общий предок ариев и этнических русских) образовался примерно 5600-5900 лет назад. Они действительно близкие родственники, в пределах нескольких столетий. Более того, при наличии древней арийской топонимики на Русском севере становится ясным, что арии вовсе не замыкались в южных степях, как полагают некоторые археологи на основе материальных признаков степной и лесостепной полосы юга России и Украины. Ясно, что арии заселяли всю Русскую равнину с юга до севера, имели практически такие же гаплотипы, как и предки современных этнических русских (с общим предком всего за несколько веков до тех времен). Поэтому совершенно разумно считать носителей гаплогруппы R1a-Z645 ариями, как и их потомков, носителей субкладов Z93-Z94-L657 и Z283-Z282-Z280. Они все современники друг друга.
Теперь посмотрим, как считают популяционные генетики. Поскольку во всех 64 37-маркерных гаплотипах (то есть на 2368 маркеров) индийских носителей гаплогруппы R1a имеется 924 мутации, то они получают, используя «скорость Животовского», 924/2368/0.00069 = 566 поколений по 25 лет на каждое, то есть 14150 лет назад. Иначе говоря, при типичных расчетах попгенетиков общий предок индийских гаплотипов группы R1a жил более 14 тысяч лет назад. Какие тогда арии? Таких ариев не бывает. Поэтому попгенетики делают следующий вывод – ариев в природе не было, а гаплогруппа R1a зародилась в Индии. Как видите, применять фантастически искаженную «популяционную скорость мутаций» вовсе не безобидно. Это тянет за собой безумные «исторические интерпретации», не имеющие ничего общего с реальностью. Более того, этому рады индийские националисты, для которых «ариев придумали англичане, чтобы порабощать индийский народ». Я понимаю, что в России эта мышиная возня националистов в Индии вообще никого не интересует, да и меня тоже, но считать нужно правильно, хотя бы во имя науки. Попгенетика и здесь показала себя лженаукой.
Взглянем еще раз на «мою» страничку в Википедии. Читаем в разделе «критика»:
Итак, автор (то есть я) так «считает» (да не только автор, а тот же Гордон Чайлдс еще в 1926 году, см. выше, о чем «критики» предпочли не упоминать). Славно, не так ли? И далее:
Я понимаю, Википедию наука не интересует, им нужно собрать все «до кучи». Но давайте разберемся. Первая статья в списке, Sanghamitra S et al. (2006), те же одиозные фамилии – Животовский, Underhill и другие. Какой метод расчета применялся? Естественно, Животовского. Какой возраст гаплотипов R1a там получили? Естественно, «exceed 10,000–15,000 years», то есть «превышающий 10-15 тысяч лет». Вывод? Никаких индоевропейцев в Индии не было. Историки и лингвисты в шоке, но не Л.А. Животовский и П.А. Андерхилл, а также под раздачу попавший бедняга Кавалли-Сфорца, член всех академий мира. Подставили его почем зря.
Следующая статья (тоже 2006) – не лучше. Группа индийцев и пара эстонцев-попгенетиков, не смыслящих в ДНК-генеалогии – Kivisild и Willems, президент эстонской академии наук, соавтор всех публикаций. Их у него, наверное, уже тысячи. Расчетов там вообще никаких нет, но есть «основополагающий» принцип – авторы полагают, что если бы арии пришли в Индию, то непременно принесли бы с собой букет разных гаплогрупп – C3, DE, J*, I, G, N, and O, а раз таких в Индии нет, то никакие арии в Индию не приходили. То, что они арии могли быть сами по себе, не смешиваться с чужаками, авторам в голову не пришло. А ведь именно это было на протяжении тысячелетий в высших кастах Индии – никаких смешиваний. Те, кто смотрели в СССР индийские фильмы в 1950-1970 гг., лицезрели индийские драмы и трагедии, как общество предохраняло высшие касты в Индии от смешивания с «низкими» кастами, и никакая межкастовая любовь не была оправданием. Так и продержались более трех тысяч лет, и сейчас гаплогруппа R1a в высших кастах достигает 72% от всех (Sharma et al, 2009). И вот эта националистическая индийская чепуха идет на «мою» страничку в Википедию как якобы критика результатов и выводов ДНК-генеалогии.
Хорошо, переходим к третьей статье (2009). Там в Википедии почти у всех авторов перепутали имена и фамилии, и Shweta Singh, например, в Википедии идет как Shweta S., как аналогично и все остальные. Ну да ладно, каков поп, в смысле, «критики», таков и приход. Что более важно, в статье опять расчеты проведены «по Животовскому». Результат – гаплогруппа R1a в Индии образовалась 13768.12 лет назад и (другой вариант) 18478.26 лет назад, если считать 25 лет на поколение, и 17623.19 и 23652.17 лет назад, если считать 32 года на поколение. Представляете, до сотой части года! Это насколько нужно быть безголовым, чтобы время до общего предка считать до сотой части года. Настоящая популяционная генетика… Здесь «метод Животовского» дает завышение в датировке в 2.75 раз и (другой вариант) в 3.70 раз. Про 32 года на поколение и говорить не приходится, это дает итоговое завышения в датировках в 3.52 и 4.73 раз. Результат – опять обсуждения того, что гаплогруппа R1a образовалась в Индии. А датировка на самом деле ниже, чем в Европе. В качестве дополнительного «аргумента» выставляется то, что «частота» гаплогруппы R1a в высших кастах Индии достигает 72%, а в европейских странах – 40%, как полагают авторы статьи. Авторы не понимают, что частота к происхождению никакого отношения не имеет. В Ирландии, например, частота гаплогруппы R1b около 92%, а гаплогруппа появилась в Южной Сибири.
Предпоследняя статья, Underhill et al. (2010, в Википедии опять ошибка), очередная «мусорная» работа в отношении расчетов, в которой изучали субклад R1a-M458. Датировка его по данным ДНК-генеалогии – примерно 4000 лет назад, образовался на границе Польши и Белоруссии. Это – западнославянский и центрально-европейский субклад, имеет две соответствующие ветви, каждой примерно по 2900 лет. Ясно, что ко времени его образования арии давно ушли из Европы на Русскую равнину, и оттуда, как носители субклада R1a-L657 – в Индию. Понятно, что эти M458 c ариями в Индию никак не могли попасть, как современные англичане никак не могли попасть в Америку с Колумбом. Но у попгенетиков свои правила, которые кроме как псевдонаукой опять же не назвать. Применяя те самые печально известные «константы Животовского», они «нашли», что возраст R1a-M458 в Польше уходит аж в мезолит, на 10700±4100 лет назад.
Эта расчетная ошибка попгенетиков во главе с Андерхиллом потащила за собой другие ошибки – они решили, что раз субклад М458 такой древний, из мезолита, то почему его нет в Индии (и в Средней Азии), куда он должен был попасть вместе с ариями? Отсюда последовал вывод, что ариев вовсе не было. То есть расчетная ошибка, бич попгенетиков, повела за собой грубую историческую ошибку.
На самом деле задачка решается в один ход – субклад М458 намного моложе субклада Z93, многочисленные потомки которого и выявлены в Индии. Практически все индийские R1a относятся к субкладу R1a-L657, возраст которого примерно 5000 лет назад (см. выше). Именно тогда в Европе только образовался субклад М458. Иначе говоря, когда R1a-L567 пришли в Индию, М458 только зарождались в Европе. Ясно, что они не могли в то же время прийти в Индию, арии из Европы давно ушли, на 500-1000 лет ранее.
К сожалению, вот таким мусором, как расчеты и исторические интерпретации в статьях попгенетиков, где есть «датировки» и соответствующие «интерпретации», заполнены академические журналы, как и цитируемый Eur. J. Human Genetics. Заметьте, там в авторах и российские попгенетики Балановские. Теперь для многих понятно, почему они «пошли свиньей» на ДНК-генеалогию, требуя ее отовсюду снять как «лженауку». Да потому что становится очевидным, что их статьи придется отзывать как ошибочные, а их, статей, – десятки, и все практически мусор в том, что касается датировок и исторических интерпретаций.
Обратите внимание, что у статьи – 34 автора, и ни один не задался вопросом, что здесь что-то не так. Вот в чем фундаментальная проблема с попгенетиками – некомпетентность и пассивность. И вот таким мусором заполнены академические издания в области попгенетики. В цитируемой статье такая обойма авторов: Underhill, Myres, Руутси, Metspalu, Животовский, King, Lin, Chow, Semino, Battaglia, Кутуев, Ярве, Chaubey, Ayub, Mohyuddin, Mehdi, Sengupta, Рогаев, Пшеничнов, Балановский, Балановская, Jeran, Augustin, Baldovic, Herrera, Thangaraj V. Singh, L. Singh, Majumder, Rudan, Primorac, Виллемс, Кивисилд. Представляете, 34 автора (российских и эстонских я выделил кириллицей, страна должна знать своих героев), и никто не понимает, что они делают, как и зачем так считают. В итоге полностью искаженное представление истории. Это же, заметим, вынесено в виде критики на «мою» страничку в Википедии, в которой делается вывод, что я неправ, и арии в Индию не приходили, и что это якобы современные представления. И дается галерея таких вот «мусорных» ссылок попгенетиков. Стыд и позор тем, кто такую ерунду написал, а редакция Википедии приняла.
Кстати, в пленарном докладе д-ра Боринской (Ин-т общей генетики РАН) на Конференции по карачаево-балкарцам, в котором половина времени (и материала) было посвящено автору этих строк, даже с показом моего портрета, она особое внимание уделила моим высказываниям из очерка «Суета попгенетиков вокруг прародителей европейцев», где я критиковал недавнюю статью («Ancient human genomes suggest three ancestral populations for present-day Europeans»), под которой подписались 98 авторов (Балановские среди них). Она в докладе представило дело так, что я якобы критиковал число авторов, мотивируя тем, что в других статьях и по 150 авторов бывает. При этом она умолчала, что на самом деле моя ключевая фраза была «я сделал критический разбор этой статьи, у которой 98 авторов, а дитя, как говорится, без глазу…». Моя критика была о том, что 98 авторов породили запутанное, вязкое исследование, изобилующее искажениями, неверными положениями, что они не состыковали геномные данные с данными ДНК-генеалогии, уже давно известными, да и просто с филогенией Y-хромосомы, которая не должна противоречить положениям и выводам статьи. Вот почему «98 авторов, а дитя без глазу». Была бы статья толковой, пусть там хоть тысяча авторов. Вот и выше – 34 автора (Underhill et al. 2010), а статья привела к неверным выводам и заключениям, которые уже несколько лет цитируют в десятках публикаций и на форумных дискуссиях, не задумываясь, что 34 автора породили мусор.
Так и со статьей с 98 авторами. Одна из выступающих с пленарным докладом на той же конференции«Этногенез, история, язык и культура карачаево-балкарского народа» (ноябрь 2014 г., РАН) объявила, что статья замечательная и продвигает наше знание в отношении происхождения европейцев. И дальше было сообщено, в чем именно продвигает – в том, что было найдено, что «происхождение древних европейцев сходится к трем древним популяциям – древние охотники-собиратели, древние фермеры, и древние евразийцы». Представляете, какой основной вывод «эпохальной» статьи 98-ми популяционных генетиков? Давайте расшифруем это замечательное высказывание. По принятым в попгенетике понятиям, «древние охотники-собиратели» – это все те, кто жили более 7 тысяч лет назад. «Древние фермеры» – это те, кто жили менее 7 тысяч лет назад. Ну, а древние евразийцы – все остальные. Переводим на нормальный язык, без этого волапюка. Вот перевод: «древние европейцы происходят от древних людей». Замечательно, не так ли? Продвигает знание. Для этого вывода понадобились 98 популяционных генетиков.
И дальше я пишу в своем очерке на Переформате:
Переходим к заключительной, пятой статье на «моей» страничке в Википедии, из серии призванных показать, что ариев в Индии не было, и что ДНК-генеалогия поэтому показывает неверные выводы, во всяком случае в отношении Индии. Из предыдущих четырех статей мы видим, какова цена этим статьям. В трех из них применен «метод Животовского» (и в двух из этих статей в авторах сам Л. Животовский), одна – просто откровенное недоразумение, где авторы рассуждают «по понятиям, а не по науке». И вот – последняя статья. Она содержит ссылки на все четыре предыдущие статьи, разобранные выше. На них статья базируется, а именно, что ариев в Индии не было, в Индию они не приходили. То есть направление статьи уже задано. Так и оказалось. Но в этой статье уже геномные данные. И что же они показывают?
Данных много, проанализировали геномы 142 человек из 30 индийских этнических популяций, а всего расматривали геномы 1310 человек из 112 популяций. Главные выводы – что Индия в целом, по популяциям, очень древний субконтинент, с высоким «геномным разнообразием». Что выявлены два основных «предковых компонента», которые характеризуются «разнообразием и частотой» – один простирается от южной и западной Азии до Кавказа, второй, который характеризуется более высоким «разнообразием», сдвинут к южной Азии, и соответствует по численности более чем 50% индийских популяций. Его древность в целом больше, чем 3500 лет, когда арии должны были прибыть в Индию. По мнению авторов, это не согласуется с «арийским вторжением» 3500 лет назад, а также ранее или позже. Правда, постоянно оговаривается, что данные по «потоку генов» сложны и неточны, что в древности таких «потоков» было много и они смазывают картину, что согласно некоторым авторам (те, что даны в Википедии и разобраны мной выше) индийцы во множестве ушли в Европу, и что даже «поток генов» между Индией и соседней Средней Азией является сложным и трудным для анализа.
Заключение статьи звучит так (убирая пустые в данном контексте фразы, например, о том, что процесс генетического структурирования в южной Азии продолжается и сейчас): «Получены сложные данные, которые трудно объяснить только предполагаемым недавним прибытием индо-ариев, но который свидетельствует о многоообразных потоках генов в южно-азиатский генетический пул, как с запада, так и с востока, на протяжении долгого времени». И дальше – про липидный метаболизм и диабет, и что нужно продолжать заниматься исследованиями. Ну и где там данные, противоречащие прибытию ариев в Индию?
Напоминаю, под каким «соусом» это помещено на «моей» страничке в Википедии. А именно, что это – «в противовес мнению иных экспертов», в смысле в противовес данным ДНК-генеалогии, и что это показывает «несостоятельность гипотез о доисторических миграциях индоевропейцев в Индию». Отается только развести руками. Пять статей «иных экспертов», три из которых откровенно неверны, так как применяли псевдонаучные «популяционные скорости», завышающие датировки на сотни процентов, одна статья полна недоразумений и построена «по понятиям», последняя вообще ничему не противоречит. Ох уж эта Википедия, которая в угоду «рецензентам», не имеющим никакой квалификации, засоряет ерундой в общем-то ценное издание.
Несколько комментариев в отношении последней статьи, по геному индийцев. На самом деле то, то авторы получили, никакого удивления не вызывает. Я уже не раз в своих статьях показывал, что, как ни странно, картина снипов в геноме следует за простенькой Y-хромосомой. Действительно, в геноме определяются сотни тысяч и миллионы снипов, а в ДНК-генеалогии Y-хромосомы для каждой гаплогруппы – всего несколько, определяющие гаплогруппу и субклад. То есть как бы хвост вертит собакой, причем хвостик совсем маленький. Любой генетик скажет – что за ерунда, мы Y-хромосому вообще почти не принимаем во внимание, когда анализируем геном. Вот в чем опять драма и трагедия современной генетики вообще и попгенетики в частности. Они не смотрят «трехмерно», за пределы своей маленькой (в этом отношении) парадигмы. А там четкие закономерности, которые генетики не видят, потому что не знают и не хотят знать филогению гаплогрупп Y-хромосомы. Связь между геномом и филогенией Y-хромосомы я описал на ряде примеров в статье «Действительно ли «генетики нашли разных русских?». Статью я закончил так: «…(надо) понять, что лежит в основе вариаций в геноме, задающих глубинные различия между популяциями. Эти различия идут из тьмы тысячелетий, и что особенно интригует – они определенно завязаны на мутации, определяющие мужские гаплогруппы. Это – совершенно новая концепция, и решать эту загадку надо в содружестве генетиков и ДНК-генеалогов».
Так вот, то, что в Индии «древнее разнообразие» – совершенно не удивительно, его диктуют, в частности (и в особенности) древние гаплогруппы L, H, J, Q, C, R2, и каждую из них можно датировать с помощью подходов ДНК-генеалогии. То, что в этом «разнообразии» тонет относительно недавняя гаплогруппа R1a в Индии – неудивительно, хотя ее около трети в современной Индии, но это только среди тех, кто может заплатить не менее 200 долларов за тест. Индийцы из джунглей в это число не входят. Поэтому вряд ли число носителей R1a в Индии превышает 15% от всего мужского населения. Это и есть в основном потомки ариев, прибывших в Индию примерно 3500 лет назад. А те, кто образует геномную связь с Афганистаном и Кавказом – это носители гаплогрупп G2a, L1b и L1c, наряду с другими, минорными. В общем, все эти геномные данные можно усилить данными ДНК-генеалогии, в том числе корректными датировками, и привести их в порядок.
Похоже, что в Индии осталось небольшое количество особенно древних носителей гаплогруппы R1a, и они иногда выявляются в джунглях, но систематических исследований их не проводилось, субклады их не определялись, в высших кастах (и в кастах вообще) они не представлены, они попадают при классификации или в низшие касты (lower caste), или в племена (tribes). Видимо, они попали в Индию в ходе древнейших миграций из Южной Сибири на запад, когда носители R1a в итоге прибыли в Европу примерно 9-10 тысяч лет назад (Klyosov, 2009-2012). Но эти данные получены на коротких гаплотипах, в которых ошибки тестирования проявляются намного больше, чем при работе с протяженными гаплотипами. А поскольку данные по этим древним R1a в Индии фрагментарные (в известном списке Sengupta [Sengupta et al., 2006] их, например, практически нет, за исключением, быть может, гаплотипов 4, 95, 109 на дереве гаплотипов на рис. 3), то систематического изучения их пока не проводилось, субклады неизвестны.
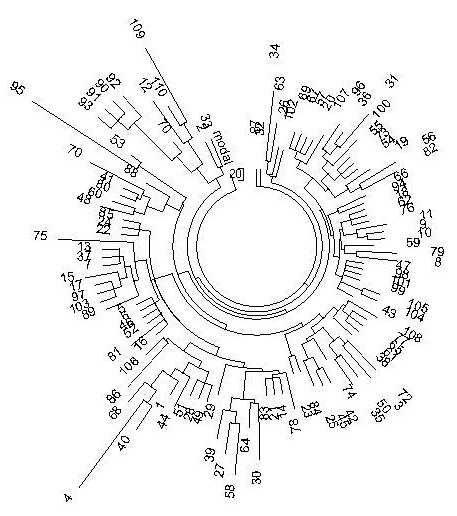
Рис. 3. Дерево из 110 10-маркерных гаплотипов гаплогруппы R1a в индийских кастах и племенах. Построено по данным Sengupta et al. (Amer. J. Human Genetics, 2006).
Формат гаплотипов – DYS 393, 390, 19, 391, 388, 439, 389-1, 393, 389-2, 461).
На данных Сенгупты (2006) стоит остановиться чуть подробнее, поскольку именно «расчеты» этих самых известных данных по Индии (более тысячи 10-маркерных гаплотипов в выборке, из них 110 гаплотипов гаплогруппы R1a) приводят популяционных генетиков к тому, что общий предок носителей R1a в Индии жил более 14 тысяч лет назад (Животовский – в авторах и статьи Sengupta et al; естественно, датировки опять рассчитывали «по Животовскому»). Дерево гаплотипов на рис. 3 не слишком симметричное, но считать вполне можно, на что указывает и полученный базовый гаплотип дерева (здесь приведены 9 маркеров, поскольку десятый, DYS461, не входит в список 37-маркерных гаплотипов, которые будут приведены ниже для сравнения, Х – маркеры, которые не определяли в работе Sengupta)
25 15 10 Х Х Х 12 10 13 11 17
Все 110 гаплотипов содержат 344 мутации, что дает 344/110/0.018 = 174 → 211 условных поколений, то есть 5275±600 лет до общего предка. Здесь 0.018 – константа скорости мутации для 9-маркерного гаплотипа, равная сумме констант для всех 9 маркеров, определенных еще в 2006 году Чандлером (Chandler, J. Genet. Geneal., 2006): 0.00076 + 0.00311 + 0.00151 + 0.00265 + 0.00022 + 0.00477 + 0.00186 + 0.00052 + 0.00242 = 0.01782 мутаций на гаплотип на поколение.
Сравним полученный базовый гаплотип с таковым для индийцев субклада R1a-L657, приведенным выше в 37-маркерном формате (общий предок жил 4750±500 лет назад):
25 16 10 11 14 12 12 10 13 11 17 – 16 9 10 11 11 24 14 20 32 12 15 15 16 – 11 12 19 23 15 16 18 19 35 39 13 11
Если переписать его в виде, удобном для сравнения
25 16 10 Х Х Х 12 10 13 11 18
то мы увидим, что базовый гаплотип, полученный по данным Сенгупты (включающих низшие и высшие касты и племена), и базовый гаплотип, полученный по данным индийского проекта FTDNA, различаются всего на одну мутацию (выделено), а на самом деле всего на 0.55 мутаций; если усреднить по всем аллелям маркера DYS19, получим DYS19 = 15.55. Другими словами, это одинаковые предковые гаплотипы, и одинаковая датировка, в пределах погрешности расчетов, безотносительно, 9-маркерные гаплотипы или 37-маркерные. Некоторое завышение датировки по данным Сенгупты, которое, впрочем, укладывается в погрешности, вызвано тремя особенно мутированными гаплотипами (под номерами 4, 95, 109, см. рис.3), на которые приходится 19 мутаций, и при их снятии получаем 325/107/0.018 = 169 → 203 условных поколений, или 5075±580 лет до общего предка.
Такова цена утверждениям попгенетиков, что ариев в Индии не было, и что общий предок индийских R1a жил более 14 тысяч лет назад. И здесь ДНК-генеалогия полностью переигрывает популяционную генетику.
Расчетный аппарат попгенетики и его фактическая лженаучность,
которая усугубляется фальшивыми «калибровками» (примеры)
В первой части этой статьи был дан иллюстративный пример расчета времени общего предка знаменитой шотландской «генеалогической» семьи МакДоналдов, которая в гаплогруппе R1a ведет свою линию от Джона Лорда Островов (John Lord of the Isles), умершего в 1386 году, то есть 628 лет назад. Было показано, что подход ДНК-генеалогии дает совершенно разумное совпадение с документальной генеалогией по Джону и его потомкам.
Документальная генеалогия дает 628 лет до годовщины смерти Джона (1386 год), а ДНК-генеалогия дает 650-700 лет до времени его жизни (1314-1364 год), то есть практически абсолютное совпадение в контексте данного исследования, учитывая, что год его рождения неизвестен.
Попробуем рассчитать те же данные «по Животовскому»:
6-маркерные гаплотипы: 17/68/6/0.00069 = 60 поколений, то есть 1500 лет до Джона.
12-маркерные гаплотипы: 44/84/12/0.00069 = 63 поколения, то есть 1575 лет до Джона.
25-маркерные гаплотипы: 109/84/25/0.00069 = 75 поколений, то есть 1875 лет до Джона.
37-маркерные гаплотипы: 178/59/37/0.00069 = 118 поколений, то есть 2950 лет до Джона.
Столь разные величины поколений и лет в результате применения «метода Животовского» получаются потому, что константы скорости мутаций для разных гаплотипов разные (самые высокие – для 37-маркерных гаплотипов), а в «методе Животовского» они для всех одинаковые – 0.00069 мутаций на маркер на 25 лет. Но это не самая большая проблема (хотя она приводит к дополнительным ошибкам почти в два раза, от 1500 лет до 2950 лет). Еще хуже то, что результаты получаются «ни в какие ворота». Не говорите об этом расчете генеалогам семейства МакДоналдов, засмеют и выгонят, откажутся разговаривать. Вот так работают «расчеты» популяционных генетиков. Как видим, разница та же, завышение в датировке на 230-450%, в данном конкретном случае. А поскольку случаи разные, то попгенетики задирают датировки обычно в 2.3-5.0 раз. Вот такая цена их расчетам.
«Знающий» попгенетик возразит – не может быть, Животовский свои скорости мутаций калибровал, это все популяционные генетики знают, и основополагающая статья его 2004 года основана на калибровках его скорости мутации, той самой, 0.00069 мутаций на 25 лет на маркер. Там всё доказано, потому его константу и взяли за основу в популяционной генетике.
И вот здесь начинается самое интересное. Похоже, ни один популяционный генетик, ни один (!), не удосужился проверить сам, лично, что там была за «калибровка». Ни один не взял исходные данные Животовского и не проверил. Я это сделал. Давайте посмотрим, как Л. Животовский калибровал. Основная его «калибровка» – по группе болгарских цыганских таборов, 8-маркерные гаплотипы; по группе полинезийцев, 10-маркерные гаплотипы; и по группе африканских банту. Идея такая – мы знаем, что цыгане появились в Европе примерно 900-1000 лет назад, в Болгарии – 700 лет назад, поэтому если взять группу их гаплотипов, посчитать мутации, приложить «популяционную» скорость 0.00069 мутаций на маркер на 25 лет, и если получится примерно 700 лет для Болгарии, или 900-1000 лет для Европы (Хорватии, например), то скорость 0.00069 – правильная. Разумно, не так ли?
Именно это Л. Животовский и объявил, что при калибровке так и получилось, именно со скоростью мутации 0.00069, так и вошло в учебники. То же самое в отношении полинезийцев и банту – у Животовского тоже получилось как надо, скорость 0.00069. А как же, калибровка-то вот она, налицо, так все уже 10 лет и повторяют. Так и ответили авторы статьи (Hammer, Животовский и др., 2009) на мои развернутые комментарии в журнале Human Genetics (Klyosov, 2009), в которых я показал, что они считают неверно, ошибаются в 3-4 раза. Авторы сослались на «the original Zhivotovsky et al. (2004) study (i.e., the effective rate was calibrated for time periods of *1,000 years). Since 2004, this approach has been employed by other researchers and critically evaluated in the scientific literature». Перевод: «Исходное исследование Животовского и др. (2004) (содержало) калибровку получающихся скоростей на протяжении примерно 1000 лет». Типа, какие могут быть вопросы, и тем более критика? Скорости калибровались, и всё тут.
Проверим, чего, похоже, никто не делал. Такая «наука» у популяционных генетиков. Давайте посмотрим для начала на цыган. Гаплотипы цыган были определены у 179 человек из двенадцати болгарских таборов. На удивление, у всех гаплотипы похожи, явно от общего не столь далекого предка. Посмотрим сначала, как надо работать с этими гаплотипами, а потом сверим, как «калибровал» Животовский. Для двойной проверки проанализируем как 6-, так и 8-маркерные гаплотипы, и если считаем правильно, результаты должны совпасть, в пределах разумных погрешностей.
У всех 179 цыган гаплотипы в 6-маркерном формате содержали 34 мутации от базового гаплотипа 15 12 22 10 11 12 (в формате DYS 19, 388, 390, 391, 392, 393). Это дает 34/179/0.0088 = 22±4 поколения до общего предка, то есть 550±100 лет. Поскольку из 179 гаплотипов 146 были идентичными друг другу, то есть базовыми, то логарифмический метод дает [ln(179/146)/0.0088] = 23±3 поколения до общего предка, то есть 575±75 лет назад. Практически полное совпадение результатов по линейному и логарифмическому методу. Это означает, что, действительно, все 179 цыган (или их подавляющее большинство) произошли от одного общего предка.
В 8-маркерном формате (дополнительно вошли DYS 389-1 и 389-2) добавилось 25 мутаций, то есть стало 59 мутаций. Получаем: 59/179/0.013 = 25±4 поколения до общего предка, то есть 625±100 лет. В 8-маркерном формате из 179 гаплотипов 126 базовые. Получаем [ln(179/126)/0.013] = 27±4 поколения до общего предка, то есть 675±100 лет назад.
Как видим, все четыре варианта расчета дали расстояние до общего предка болгарских цыган одно и то же, в пределах погрешностей расчета: 550±100, 575±75, 625±100 и 675±100 лет назад. Надо отметить, что хотя присутствие цыган в Болгарии было отмечено 700 лет назад, но эти сведения не обязательно точны, они могут относиться к другим цыганам, не обязательно гаплогруппы Н1 (как в данном случае), потомство у первых цыган могло не выжить, и так далее. Тем не менее, 550-675 лет назад – это не слишком далеко от отмеченного времени «начала присутствия» цыган в Болгарии.
Читатель, видимо, уже приготовился, что у Животовского получилось значительно больше, то есть не менее 1500 лет до общего предка цыган, и что он повинился, что такого науке не известно, значит, ошибочка вышла с константой 0.00069. Слишком занижена. Ведь цыган было 179, мутаций (в 8-маркерных гаплотипах) 59, так что у Животовского должно получиться 59/179/8/0.00069 = 60 поколений, то есть 1500 лет до общего предка. Но наиболее понимающие читатели, зная популяционных генетиков, уже заулыбались. Потому что у тех раз результат заранее известен, такой и получится. Наука такая. Надо просто уметь рукава пониже держать, желательно, чтобы руки закрывали.
Следим за рукавами. Один гаплотип был из списка удален, потому что он «оказался нечетным» (?). Какой удалять – естественно, на усмотрение удаляющих (или базовый, или мутированный, то есть уже направленно вносится искажение серии данных). Далее постулировали, что общий предок всех цыган жил 700 лет, то есть 700/25 = 28 поколений назад. Замечаете прием? Я-то выше рассчитывал, когда жил общий предок (получилось в среднем 600 лет назад), а Л. Животовский просто постулировал, что 700 лет назад, и отсюда вел отсчет. Но есть большая разница – время, когда цыгане были замечены в Болгарии, и время, когда жил общий предок рассматриваемых 179 цыган. Совсем не обязательно, что он был первым цыганом в Болгарии, как уже отмечено выше.
Но 600 и 700 лет – разница не так велика, так как же он подтвердил свою скорость мутации в три раза ниже?! А вот как – после совершенно произвольного снятия одного гаплотипа получилось, что при использовании квадратичного метода среднее число мутаций на маркер было 0.01272, и при делении на постулированные 28 поколений до общего предка средняя скорость мутации на маркер оказалась 0.01272/28 = 0.000454. А надо 0.00069, непорядок. Надо данные менять, иначе «теория» под угрозой. Животовский с коллегами написали в статье, что это была «недооценка». Поэтому решили пересчитать данные, введя некие «веса», и в ходе этого сняли еще ряд гаплотипов цыган. После каждой стадии подгонки смотрели, насколько скорость удалось подогнать к 0.00069, но никак не получалось. Тогда решили, что один табор – «Музыканты» – «очень гетерогенный» (на самом деле, по моим расчетам, он был совершенно нормальный, как показано выше при сопоставлении линейных и логарифмических методов), и сняли его целиком, все 19 гаплотипов, и опять ввели «веса по размеру популяции» (что ни в коем случае нельзя делать). Получили среднюю скорость мутации 0.000725±0.000187 (обратим внимание на величину погрешности), и решили, что это и есть 0.00069.
Если это «калибровка», то непонятно, что такое калибровка в понимании авторов «основополагающей» статьи, на которую все попгенетики обязаны ссылаться, иначе рецензенты статью в журнал не пропускают. На это жаловался венгерский автор на форуме по «генетической генеалогии», о чем я уже писал на Переформате. Вот такая популяционная генетика. На самом деле, авторы зря занимались акробатикой. Раз надо получить 0.00069, то способ простой – надо было повыкидывать гаплотипы с мутациями как «гетерогенные» или просто дефектные, или те, которые просто не понравились, оставить на оставшиеся полторы сотни человек ровно 23 мутации, и получить 23/150/8/0.00069 = 28 желаемых поколений, то есть как раз искомые 700 лет. «Калибровка» была бы не хуже, и никаких хлопот. Как говорил хрестоматийный герой – «как пожелаем, так и сделаем».
Так же рассчитывали и по гаплотипам полинезийцев (маори, самоанцы и жители островов Кука). Решили исходить из того, что маори прибыли в Новую Зеландию 800 лет назад, то есть 32 поколения назад, по 25 лет на поколение. Гаплотипы были 10-маркерные, но акробатика пошла и там. Самоанцев решили в расчет не включать, а у остальных сократить гаплотипы до 7-маркерных. Видимо, с 10-маркерными не получалось то, что «пожелали». Применяя квадратичный метод, нашли, что среднее число мутаций на маркер в 7-маркерных гаплотипах у маори и островитян Кука равно 0.00998, и разделив на 32 поколения, получили таким образом «форсированную» (в отличие от естественной) скорость мутации 0.00032. Как видно, это вовсе не 0.00069, а в два с лишним раза медленнее. Авторы заключили, что это опять «недооценка», и снова пустились в долгие и вязкие рассуждения, почему это может быть недооценкой. На этом «калибровка» данной серии гаплотипов была завершена.
С африканскими банту и того хуже. Раздел «Материалы и методы». Смотрим про африканских Банту. Читаем – «анализировали 148 человек гаплогруппы Е3а7-М191», перечисляются племена, сообщается, что анализ проводили по 10-маркерным гаплотипам.
Раздел «Результаты». Данных по Банту нет, они вообще в данном разделе не упоминаются.
Раздел «Обсуждение». Банту не упоминаются.
Раздел «Применение». Говорится, что применили полученную величину 0.00069 к популяции Банту, чтобы посмотреть, как это коррелирует с археологическими, лингвистическими и историческими данными. Далее говорится, что если усреднить по всем 11 племенам и популяциям, перечисленным в разделе «Материалы и методы», то получится 3400±1100 лет до «экспансии» Банту в подгруппе со снипом М191. И далее – «археологические и лингвистические данные поддерживают это значение, поскольку показывают присутствие Банту в Западной Африке в неолитические времена ∼1000 лет до нашей эры, или даже 2000 лет до нашей эры». И далее – «Величина 3400 лет назад может рассматриваться как нижняя граница для времен экспансии Банту… Если мы возьмем наиболее частые гаплотипы среди тех 148 хромосом как предковые, и приложим к ним величину 0.00069, то время для М191 получится 14700 лет… В этом случае, экспансия Банту могла произойти ранее, чем 3500 лет назад». И далее идут долгие рассуждения с неопределенной концовкой. Остается совершенно загадочным, причем здесь 14700 лет назад, когда усреднение дало 3400 лет назад «до экспансии», и причем здесь вообще «экспансия». Иначе говоря, авторы просто перешли на другую тему, и про «калибровку» успешно забыли. Нет её там.
Если это «калибровка», то что такое не калибровка? К сожалению, исходные данные (гаплотипы) в статье не приведены, ссылка статьи, что они «в сети», не дает никакого линка. Я могу только заключить, что рассматриваемая популяция Банту молодая, не более 1000-1200 лет до общего предка, как это часто бывает с африканскими популяциями, только недавно прошедшими очередное бутылочное горлышко. Ни к каким археологическим или лингвистическим данным это не имеет никакого отношения. Рассуждения в статье не имеют никакого значения в отношении рассматриваемого вопроса. Никакой «калибровки» не было и близко.
Такова цена «калибровкам» Л.А. Животовского, которые стали краеугольным камнем популяционной генетики в последние 10 лет. Кроме как «стыд и позор» других слов я не подберу. Последние пять лет я объясняю это в научной литературе, начиная со статьи в Journal of Genetic Genealogy (2009) и Human Genetics (2009), и далее в журнале Advances in Anthropology, выступаю на англоязычных форумах. Но попгенетикам – как о стену горох. Они продолжают плодить в академической литературе мусор. А те, кто в этом ровным счетом ничего не понимают, как тот же воронежский «индолог» Семененко, тем не менее, вылезают со своим критическим словом, что «метод Животовского» самый правильный, потому что им пользуются все популяционные генетики в мире. И далее – «Я опираюсь на данные международных коллективов профессиональных генетиков, опубликованные в ведущих международных научных журналах по генетике». Грош цена этим «международным коллективам профессиональных генетиков», а также рецензентам этих «международных научных журналов», а также неучам, кто это бездумно повторяет.
Напомним, что Л. Животовский применил совершенно усредненные и искусственные приближения, приняв, что константа скорости мутаций в Y-хромосомах ДНК должна быть усреднена по 10 тысячам (!) гаплогрупп за все время существования человечества, и тогда у него получилось, что она, константа скорости мутации, должна быть равна 0.00069 мутациям за 25 лет на маркер, причем она одинакова для любых гаплотипов, 6-, 7-, 8-, 9-, 10-, 12-маркерных и так далее. Любой, кто имеет хотя бы минимальный опыт работы с гаплотипами знает, что для каждого маркера есть своя константа скорости мутации, и в 6-маркерные гаплотипы попадают одни маркеры, в 8- или 9-маркерные свои (и, значит, средняя константа скорости мутации будет уже другой), в 10- или 12-маркерные гаплотипы добавляются еще другие маркеры со своими константами, и так далее. Считать, что константа одна на все – это, во-первых, принципиально неверно, во-вторых – это вносить значительную погрешность в расчеты. Именно эту ошибку делают все без исключения «международные коллективы профессиональных генетиков», чьи фундаментально ошибочные статьи «опубликованы в ведущих международных научных журналах по генетике».
Это – трагедия современной популяционной генетики. А люди, которые в этом совершенно не разбираются, продолжают кудахтать, другого слова не подберу, что «он не генетик» (это про меня). Но генетика здесь совершенно не при чем, нет в ДНК-генеалогии генетики, генетика закончилась до ДНК-генеалогии, когда генетики определили, какие мутации есть в ДНК, сколько «тандемных повторов» есть в маркерах, и какие снипы найдены в каких участках Y-хромосомы. Иначе говоря, генетика, а точнее, лаборанты в компаниях типа FTDNA или в лаборатории популяционной генетики РАМН, снабжают нас этими данными, а ДНК-генеалогия идет дальше, проводя количественную обработку этих данных, чего генетики делать уже не умеют. У них образование не то, их этому не учили. А учиться, как показывают последние 6-8 лет, они либо не могут, либо не хотят.
Что же касается того, что наши с И.Л. Рожанским калибровки констант скоростей мутаций «не дали ничего нового», и «просто повторили давно известное» (то, что заявил Балановский с трибуны в ходе своего доклада), то это только указывает на уровень компетенции «специалиста». Уровень Балановского – ужасающе низкий. Совершенно серый уровень, никакой творческой искры, сплошные штампы.
Кстати, у меня на конференции был продолжительный разговор с Л.А. Животовским, в ходе которого был снят целый ряд недоразумений, как-то застрявших в голове Льва Анатольевича. Он выставил мне четыре вопроса (они-то и были недоразумениями), и остался ответами полностью удовлетворен, вопросы снял. Один из них отражал традиционное заблуждение (скорее, дезинформацию) попгенетиков, что я якобы приравниваю гаплогруппу к этносу. Как только я напомнил, что везде пишу, что у этнических русских есть четыре основных гаплогруппы и полтора десятка минорных, у карачаево-балкарцев три основных гаплогруппы и целый ряд минорных, он тут же сказал – понял, вопрос снимаю.
Другой вопрос был о том, почему я не учитываю «пропадания» генеалогических линий, поскольку по причине этого он и использует сильно заниженную константу скорости мутации (0.00069, а не 0.00179 для 67-маркерных, или 0.00243 для 37-маркерных), чтобы компенсировать потери. И еще, почему я не учитываю размер популяции, поскольку когда у отца рождается в итоге 40 сыновей, а у тех еще по много сыновей, то идет быстрый разбег мутаций в ДНК сыновей и прочих внуков и правнуков, и это надо учитывать. Об этом он рассказывал в своем докладе.
Я пояснил, что это принципиальное заблуждение популяционных генетиков, потому что когда, например, между общим предком и мной 40 поколений, то мне все равно, сколько у меня братьев и прочих родственников, поскольку мутации между общим предком и мной идут исключительно на протяжении этих 40 поколений, только по этой вертикали, по этой генеалогической ниточке. У остальных выживших родственников от того же общего предка – то же самое, те же 40 поколений, как и у всех остальных. Поэтому у всех нас сегодня, у каждого потомка этого общего предка, в 111-маркерных гаплотипах в среднем по 8 мутаций. У одних 7 мутаций, у других 9 мутаций, но в среднем 8 мутаций у каждого. Если нас, протестированных, собрать 100 потомков от этого общего предка, то на всех придется 800 мутаций в наших 111-маркерных гаплотипах. Вот и получаем: 800/100/0.198 = 40 поколений до общего предка (я здесь для простоты пренебрегаю возвратными мутациями). А у тысячи потомков будет 8000 мутаций, и получим 8000/1000/0.198 = 40 поколений, то же самое. А у 20 потомков будет 160 мутаций, и получим 160/20/0.198 = 40 поколений, то же самое. И это не зависит, нас только 20 человек выжило в потомках, или 100, или 1000 человек (просто погрешность расчетов больше у малого числа гаплотипов). Потому что мутации в каждом из нас индивидуальны. Здесь, кстати, 0.198 – это константа скорости мутации в 111-маркерных гаплотипах.
Лев Анатольевич долго мучился, пытаясь побороть в себе это старое заблуждение попгенетиков, и я, чтобы ему помочь, привел пример с радиоактивным распадом. Там неважно, образец радиоактивного материала в килограмм весом, или в грамм, или в миллиграмм. Время полураспада будет одинаковым, и, значит, и константа скорости распада будет той же самой. Процесс-то первого порядка, как и мутации в гаплотипах. По такому же принципу размер образца (число потомков) не влияет на константу скорости мутации, и у попгенетиков – принципиальное заблуждение на этот счет. В итоге Л.А. согласился, но сказал, что еще помоделирует, что я приветствовал. Мы даже условно договорились написать совместную статью, в которой разобрать и сравнить основные принципы и положения популяционной генетики и ДНК-генеалогии, но позже Л.А. Животовский от такой статьи отказался. Нетрудно догадаться, почему.
Правда, несколько позже выяснилось и другое. В первой части этой статьи я уже писал, что на Конференции была зачитана (профессором Э.К. Хуснутдиновой) Резолюция Круглого стола по антропологии, генетике и ДНК-генеалогии, в которой не было никаких слов «осуждения» ДНК-генеалогии, да их и не могло быть, поскольку аудитория принимала ДНК-генеалогию с энтузиазмом, и даже изгнала из зала Балановских, которые пытались устроить обструкцию дополнительной лекции по ДНК-генеалогии, на которую собрался зал. Иначе говоря, был провал противников ДНК-генеалогии.
Однако Балановская изготовила подметную «резолюцию», и разместила ее еще 27 ноября на сайте своей лаборатории популяционной генетики РАМН, то есть именно в день своего изгнания, когда для Конференции оставался еще полный день работы. Далее она начала проталкивать этот подметный вариант, который на Конференции не звучал, через Оргкомитет Конференции и докладчиков. Концовка его следующая: «в рамках распространяемой в интернете «ДНК-генеалогии» отождествляются принципиально разные этнические и генетические категории, прикрепляются этнические названия к генетическим «родам» и археологическим культурам. Понятийно-расчётный аппарат ДНК-генеалогии научно необоснован, не прошёл научную апробацию в индексируемых научных журналах и основан на смешении популяционно-генетических, генеалогических и историко-культурных понятий. Все эти черты ДНК-генеалогии выводят ее за рамки академических наук и находятся в противоречии с необходимостью корректного подхода, различающего биологические данные о популяции от понятия народа во всей его сложности. Поэтому упоминание ДНК-генеалогии в названии Круглого стола неуместно».
Спустя 10 дней, Балановская прислала эту «резолюцию» мне для «согласования», тем самым фактически подтвердив, что на ее сайте была размещена фальшивка, а не согласованная Резолюция. При этом она сообщила, что «Проект резолюции разработан учеными, представляющими и генетику, и антропологию: Е.В. Балановской, О.П. Балановским, Л.А. Животовским, Д.В. Пежемским. Также текст был одобрен В.А. Шнирельманом, мнение которого особенно важно в плане формулировок, касающихся этносов. Окончательные формулировки были утверждены председательствовавшим модератором (Н.К. Янковским)».
Характерно, что фамилий профессора Э.К. Хуснутдиновой и профессора М.Д. Каракетова, основных модераторов Круглого стола, среди перечисленных «разработчиков» нет. Пикантно и то, что именно Э.К. Хуснутдинова зачитывала настоящую Резолюцию на заключительном пленарном заседании Конференции, но не попала в список «разработчиков». Как и М.Д. Каракетов, который пытался остановить Е. Балановскую в ходе ее доклада, когда она давала мне персональные (негативные, разумеется) характеристики, но остановить ему не удалось. Балановская его откровенно проигнорировала, и, как видно, к составу «разработчиков резолюции» не подпустила. Зато там оказался В.А. Шнирельман, которого не было ни среди модераторов, ни среди докладчиков Круглого стола, и который, уверяю, не имеет никакого понятия о ДНК-генеалогии.
Кстате о В. Шнирельмане. Совершенно очевидно, что его привлекли к «одобрению» не случайно. Он – автор печально известного доклада при Московском бюро по правам человека, доклада под названием «Мифы современного расизма в РФ», в котором он как пугало выставляет древних ариев, раздел доклада называется «Праиндоевропейцы и русский национализм», и весь доклад построен в издевательских конструкциях в отношении изучения древней истории русского народа. «Расисты» для Шнирельмана – это те, кто занимается древней историей славян, их он называет «идеологами», и они, конечно, Шнирельману неприятны. Он пишет:
Разумеется, это Шнирельману крайне неприятно. В список таких «идеологов», осуждаемых Шнирельманом, попали известный востоковед И.М. Дьяконов и лингвисты В.В. Иванов и Т.В. Гамкрелидзе, а также «мифотворец-патриот Л.Н. Гумилев» (о, патриот – это бранное слово у Шнирельмана), сибирский археолог В.Е. Ларичев («пропагандировались фантазии сибирского археолога В.Е. Ларичева о древнейшей в мире цивилизации в Сибири, созданной, естественно, индоевропейцами»), писатель В.А. Чивилихин (который «настаивал на том, что предки славян были автохтонами в поволжских и причерноморских степях. В итоге он договаривался до того, что славяне будто бы существовали как общность уже пять тысяч лет назад», и «не без удовольствия замечал, что и в долине Хуанхэ древнейшее население было представлено светлокожими индоевропейцами» (какое уж там удовольствие для Шнирельмана…).
В число «идеологов» попали также академик Б.А. Рыбаков, в трудах которого «рисовалась многотысячелетняя история первобытных славян» (это – крайне неприятно для Шнирельмана), как и то, что «корни славян Рыбаков издавна искал в бронзовом веке», и «Рыбаков делал все, что в его силах, чтобы обнаружить славянские корни в глубинах первобытности». Шнирельману крайне неприятно то, что академик Рыбаков «полагал, что доказательство глубокой древности славян положительно скажется на самосознании и самоощущении русского народа. Не случайно одну из своих популярных статей он назвал «глубокие корни – могучая крона»». С точки зрения Шнирельмана, это плохо. Это – плохая «идеология». Это – «национализм». Интересно, с каких пор шнирельманы диктуют нам, русским, что такое «национализм», и почему русский патриотизм – это плохо, это плохая «идеология».
В число «идеологов» попал и О.Н. Трубачев, который, по мнению Шнирельмана, является «важным источником информации для русских националистов», а именно его «теория… о близком родстве и теснейших контактах между славянами и индоариями в Северном Причерноморье». И далее – «…Трубачев всеми силами пытался доказать, что после ухода оттуда основной массы их соплеменников в Переднюю Азию какие-то группы индоариев надолго задержались в Северном Причерноморье и вполне могли иметь тесные контакты с ранними славянами». Это «не осталось незамеченным в стане русских националистов». «В своих псевдонаучных построениях они …следовали разработкам Иванова и Гамкрелидзе», а другие «вновь вернулись к идее балканской прародины». По мнению Шнирельмана, это очень плохо, ну, не могут древние русские и прочие славяне быть «щедрым народом, отважными воинами и неутомимыми завоевателями», это все вредные мифы. А уж арии – это пунктик у Шнирельмана. Арии непременно относятся к нацистской Германии. И так далее, полсотни страниц вот такого ненавистного по отношению к истории славян текста. Может, это и есть расизм, только шнирельмановский?
Надо сказать, что за прошедшие со времени его доклада несколько лет основные его пассажи в отношении отсутствия древней истории славян оказались полностью отвергнуты если не «официальной наукой», то научными данными, в первую очередь ДНК-генеалогией. Подтвердились выводы и В.Е. Ларичева, и И.М. Дьякова, и Б.А. Рыбакова, и О.Н. Трубачева, и В.В. Иванова и Т.В. Гамкрелидзе, и Л.Н. Гумилева, и В.А. Чивилихина, но никак не Шнирельмана.
Понятно, что он с готовностью принял приглашение Балановской, для того, чтобы «текст был одобрен В.А. Шнирельманом, мнение которого особенно важно». Ну, понятно, не бином Ньютона. Для Балановской и Шнирельмана древность славян – как нож по сердцу. Поэтому надо «валить и мочить» ДНК-генеалогию. Вот так делаются дела в РАН, и наглость тех, кто так «делает», не имеет предела. История опять повторяется, в виде фарса.
Так вот, я написал Л. Животовскому, и спросил, действительно ли он был среди «разработчиков» этой «резолюции». Он ответил, что проект резолюции ему прислали в начале декабря (то есть когда этот «проект» уже неделю подавался Балановскими в СМИ как якобы принятый Конференцией, то есть окончательный), и по сообщению Л. Животовского, он, то есть Животовский, вычеркнул из «проекта» слова «выводят ее (т.е. ДНК-генеалогию) за рамки академических наук», а все остальное оставил. То есть оставил и «в рамках распространяемой в интернете «ДНК-генеалогии»», и «прикрепляются этнические названия к генетическим «родам» и археологическим культурам», и «понятийно-расчётный аппарат ДНК-генеалогии научно необоснован», и «не прошёл научную апробацию» и «основан на смешении популяционно-генетических, генеалогических и историко-культурных понятий», и «находятся в противоречии с необходимостью корректного подхода».
И это после того, как он снял практически все свои вопросы в ходе нашей с ним дружеской дискуссии в очереди в академической столовой и за самим обедом. Все вопросы снял! А если не снял – имел все возможности снять. Он же знает прекрасно про мои статьи и книги в области ДНК-генеалогии, но поддерживает слова «распространяемой в интернете»! Он прекрасно знает, что я не прикрепляю никаких «этнических названий» к археологическим культурам, да и как это возможно? У них уже есть названия. Скажем, майкопская культура – как можно к ней применять «этническое название»? – эти названия все молодые, а культура – старая. Как Л. Животовский может поддерживать «положение» о том, что расчетный аппарат ДНК-генеалогии «научно необоснован»? Что он «находится в противоречии с необходимостью корректного подхода»? Все это Животовский прекрасно знает, но… Помните, что Ленин говорил про интеллигенцию, что это вовсе не мозг нации? Вот то-то и оно. Это он и про Животовского говорил. На меня, признаться, Л. Животовский при личном общении поначалу произвел хорошее впечатление. Видимо, по сравнению с Балановской, которая произвела при личном общении впечатление крайне отталкивающее. Но, увы, с Животовским произошла ошибка. Манеры забили сущность. Нерукопожатность не выявилась. Поначалу.
Возвращаемся к «методу Животовского» и «генеалогическому методу» в исполнении попгенетиков. Первый – вообще чудовищно неправилен в любом исполнении, второй в исполнении попгенетиков просто примитивен, и потому неправилен. Там не делят серии гаплотипов на ветви, там смешивают общих предков ДНК-линий, там получают фантомные датировки, в общем, все равно получается ерунда. Но поскольку Балановские думать не умеют, то они в последнее время на всякий случай используют оба метода, так в статьях и описывают – «метод Животовского», и «генеалогический метод». А потом подбирают, какой им лучше подойдет, по обстановке. Это что, наука? Позор это, а не наука. Разобраться они так и не смогли, и, естественно, их датировки в таком бессильном варианте никому не нужны. Разница их – обычно в три раза друг с другом. Вот и судите об уровне их компетенции и «творческой» пассивности.
Критика попгенетики со стороны ДНК-генеалогии в печати
В 2009 году в журнале Human Genetics вышла статья группы авторов – Michael Hammer, Doron Behar, Tatiana Karafet, Fernando Mendez, Brian Hallmark, Tamar Erez, Lev Zhivotovsky, Saharon Rosset, Karl Skorecki – по датировке общего предка коэнов (по Библии и Торе – потомков Аарона, брата Моисея, которого (Аарона) Всевышний назначил быть первосвященником. Эта работа была мной задолго до того выполнена, и я показал, что общий предок коэнов (гаплогруппы J1) жил чуть больше тысячи лет назад (1,075±130 лет назад – линейным методом, и примерно 1025 лет назад – логарифмическим). Понятно, что по датировке нынешние коэны этой гаплогруппы никакого отношения к Аарону не имеют, поскольку библейский Аарон должен был жить примерно 3600 лет назад, во времена библейского же исхода из Египта. Мои данные показали, что, возможно, их предки прошли «бутылочное горлышко популяции» и продолжили свою ДНК-линию в 10-м веке нашей эры. Но, по предположению Президента международной ассоциации коэнов в нашей с ним личной переписке, возможно, тот уважаемый еврей тысячу лет назад был просто назначен коэном, или избран местной еврейской общиной, поскольку общине был нужен легитимный лидер, а наследственного коэна, чтобы продолжить дело, не оказалось. Такая процедура предусмотрена иудейским религиозным законодательством. Поэтому общий предок нынешних коэнов и жил примерно тысячу лет назад, в 10-м веке нашей эры плюс-минус сотня лет.
Но указанная группа авторов (Hammer, Животовский и др.) применили, разумеется, «скорость Животовского», и получили, разумеется, «время жизни Аарона» в библейские времена, а именно 3190±1090 лет назад, о чем торжественно заявили, что получили прекрасное совпадение с ожидаемым результатом. Статья закачивается так: «Полученная датировка согласуется с гипотезой, что «линия коэнов» является уникальной ДНК-линией древних евреев, и унаследуется по отцовской линии вместе с линией еврейского духовенства». Там же обязательные фразы, что история еврейских священников сложна, и ее необходимо продолжать изучать.
Поскольку результаты и выводы статьи были фундаментально ошибочными, я написал откровенно разгромную статью в тот же журнал, Human Genetics, на 14 страницах, в которой подверг резкой критике «метод Животовского» и полученные результаты, и пересчитал заново все их данные, приведя совершенно красноречивые деревья гаплотипов. Статья была принята к печати через неделю после ее получения редакцией. Ответ попгенетиков ждали больше месяца, задержав выпуск журнала. Ответ пришел совершенно беззубый, вялый и неконструктивный. И понятно почему – крыть авторам было нечем, можно было только вяло возражать, не приводя никаких данных или разумных аргументов. «Аргументы» были следующими, в сумме их было восемь:
1. Оппонент (то есть я) выступил с критикой слишком быстро, что «не есть в интересах основательного научного процесса».
2. В свой критике я ссылаюсь на недоступные и неопубликованные работы (в печати в J. Genetic Genealogy).
3. Использую нестандартную терминологию, как, например, «логарифмический» и «линейный» методы.
4. Я сделал неверный вывод, что гаплотипы образуют «генеалогию» и к ним могут применяться «генеалогические» подходы (то есть не «популяционные»).
5. Я не учитывал «генетический дрейф» или «никакие другие особенности мутаций в микросателлитах».
6. Авторы не знают, насколько мой метод применим к их системе.
7. Применяемый метод Животовского правильный, откалиброван и критически рассмотрен в научной печати.
8. Применяемые мной «деревья гаплотипов» являются интересными и могут быть в высшей степени полезными, но к ним нужно относиться с «большой осторожностью».
Положение (1), конечно, вызывает смех. Авторы, видимо, не в курсе, что «метод Животовского» уже давно вызывает насмешки, что я его давно и детально разобрал (что и делаю опять в данной статье), и на это описание не нужно времени.
Положение (2) можно было бы принять, если бы не русскоязычные Л. Животовский и Т. Карафет в авторах. Один вопрос мне в частной переписке, если они не умеют пользоваться Интернетом, и я бы дал им ссылки на работы, на которые они легко могли бы взглянуть. Мог бы и выслать. Если бы они хотели. Но желания, явно, не было.
Положение (3) – авторы просто не владеют предметом. Логарифмический метод в химической кинетике существует уже десятки лет. Он подробно и в азбучном варианте изложен в учебнике И.В. Березина и А.А. Клёсова «Практический курс химической и ферментативной кинетики» (1976), который уже почти 40 лет служит основным учебником по данному предмету для университетов. Опять – короткий вопрос со стороны Л. Животовского или Т. Карафет, и они узнали бы для себя много нового. Но не захотели.
Положение (4) – авторы опять не поняли, о чем речь, или сделали вывод, что не поняли. А речь о том, что дерево гаплотипов можно разложить по ветвям, и к каждой ветви приложить «генеалогический подход» и «генеалогическую скорость мутаций». А вот обрабатывать все дерево чохом, поперек ветвей, как делают авторы, категорически неверно.
Положение (5) – авторы снова не поняли, что разложение дерева по ветвям – это и есть учет «генетического дрейфа». Фраза «Никакие другие особенности мутаций в микросателлитах» может только вызвать смех. Какие «другие особенности» – авторы не пояснили.
Положение (6) – в моем «Комментарии» всё описано. Авторы просто не захотели, или не смогли разобраться.
Положение (7) – неверно и подтасовочно. Мы уже знаем, какой он «правильный», как «откалиброван» и как «критически рассмотрен в научной печати».
Положение (8) – наконец-то, разумные слова. Следует, правда отметить, что к «методу Животовского» слова про «осторожность», по мнению авторов, видимо, неприменимы. Там осторожность не нужна. Это только к другим авторам относится. Во всяком случае, прошло более пяти лет, и слова «являются интересными и могут быть в высшей степени полезными» так и остались словами. Судя по сегодняшней реакции Балановских, эти слова они к действию не приняли. Напротив, ДНК-генеалогия у них вызывает озлобление, и я уже пояснил, почему. Если в 2009 году они еще были начинающими, то сейчас погрязли в неверных данных, статьях и интерпретациях, пройдя точку невозврата.
С тех пор я многократно критиковал «метод Животовского» в печати – и в Вестнике Академии ДНК-генеалогии, и в J. Genet. Geneal., и в книге «Происхождение славян», в главе 16, и в журнале Advances in Anthropology. Этот «метод» неоднократно критиковал Dienekes Pontikos в своем блоге, подчеркивая его фундаментальную неприменимость для расчетов по сериям гаплотипов. Но попгенетики литературу не читают и литературы не знают, кроме своих собственных статей.
Отсутствие критики ДНК-генеалогии
со стороны попгенетики в научной печати
Этот раздел исключительно краткий. Просто потому, что в научной печати никакой критики ДНК-генеалогии не было. Вся «критика» сводится к совершенно неквалифицированным, а зачастую злобным комментариям в сети (Балановские, Дробышевский, Семененко и многие другие «любители»). То, что на ноябрьской Конференции в РАН в своих докладах говорили Боринская и Балановские, это не критика, это набор в лучшем случае недоразумений и искажений, в худшем – откровенная ложь. Об этом – в следующих частях этой статьи.
Продолжение следует…
Анатолий А. Клёсов,
доктор химических наук, профессор
Перейти к авторской колонке
Профессор А.А. Клёсов после удачной охоты
Продолжим сравнительное рассмотрение ДНК-генеалогии и популяционной генетики, их расчетные методы и характер выводов. Рассмотрим пример, имеющий важное историческое значение. Он имеет прямое отношение к легендарным ариям, которые по историческим сведениям (скорее, предположениям) перешли в Индию примерно 3500 лет назад. Это впервые было высказано, видимо, немецко-английским ученым-ориенталистом Максом Мюллером (1823-1900), и эта дата приводится в книге Гордона Чайлдса «Арии. Основатели европейской цивилизации» (1926). Он пишет: «…Ригведа была создана вскоре после 1400 года до н.э., и вторжение ариев в Индию относится примерно к тому же времени» (то есть примерно 3400 лет назад). Но в следующем абзаце Чайлдс продолжает – «Недавно вызов этой традиционной точке зрения был брошен сразу с двух сторон. Паргитер считает, что проникновение ариев в Индию началось задолго до составления ведических гимнов… Изучение генеалогий правителей привело упомянутого автора к заключению, что арии… вторглись в Индию скорее ближе к началу, чем к концу 2-го тысячелетия до н.э. …Однако в последние годы она была оспорена и с другой стороны. Брюннхофер и некоторые другие исследователи утверждают, что …проникновение ариев в Индию следует относить к гораздо более позднему времени, чем это обычно предполагается».
Итак, имеем датировки для перехода ариев в Индию – примерно 3400 лет назад, примерно 4000 лет назад, и позже, скажем, 3000-2800 лет назад. Такой разброс и остался с 1926 года до настоящего времени. Поскольку четкой археологии, которая дала бы уточнение датировок, нет, то историческая наука так и осталась в отношении ариев в Индии на том же уровне, что и 90 лет назад. Индийский эпос в основном иносказательный, из него для исторической науки много не получить. Древние названия местностей в Индии, как Арьяварта, датировкам не помогают. Поэтому развелось много ревизионистов-«псевдоученых», так называемых «индологов», которые несут такое, что уши вянут. Например, что ариев вообще не было, или что они, напротив, вышли из Индии и пришли в Европу. Всё это утверждает, например, некто А. Семененко из Воронежа, опять же «индолог». Его в ярость, сравнимой с таковой у Балановских, приводят данные ДНК-генеалогии с датировками общих предков у исторических ариев.
Давайте посмотрим на эти данные, и сравним, что говорит об ариях в Индии ДНК-генеалогия, и что говорит попгенетика. Мнение некого А. Семененко мы отбрасываем как псевдонаучное и не заслуживающее внимания. Когда он поймет, вдумается, сам с покаянной придет, тогда и рассмотрим его «мнение».
В сети есть Проект Индии (FTDNA), это фактически база данных индийских гаплотипов. В ней 187 гаплотипов в 37-маркерном формате, из них 64 гаплотипа гаплогруппы R1a, то есть 34% от всех. Дерево всех 187 гаплотипов выглядит следующим образом (построено с помощью профессиональной компьютерной программы PHYLIP 3.695 с преобразованием в программе MEGA6).
Давайте посмотрим на эти данные, и сравним, что говорит об ариях в Индии ДНК-генеалогия, и что говорит попгенетика. Мнение некого А. Семененко мы отбрасываем как псевдонаучное и не заслуживающее внимания. Когда он поймет, вдумается, сам с покаянной придет, тогда и рассмотрим его «мнение».
В сети есть Проект Индии (FTDNA), это фактически база данных индийских гаплотипов. В ней 187 гаплотипов в 37-маркерном формате, из них 64 гаплотипа гаплогруппы R1a, то есть 34% от всех. Дерево всех 187 гаплотипов выглядит следующим образом (построено с помощью профессиональной компьютерной программы PHYLIP 3.695 с преобразованием в программе MEGA6).
Рис. 1. Дерево 187 индийских гаплотипов Y-хромосомы в 37-маркерном формате. Слева – плотная ветвь из 64 гаплотипов гаплогруппы/субклада R1a-L657, с общим предком, жившим 4750±500 лет назад; справа и внизу – серии древних индийских гаплотипов гаплогрупп (по часовой стрелке) Н, L, Q, L, J-M304, O, C, L, C, J1-M267, J2b-L282, J2-M172, R2-M124, Q (повторы гаплогрупп – это отдельные ветви этих гаплогрупп). Общие предки ветвей справа и внизу жили 8-12 тысяч лет назад.
Построено по данным Индийского проекта FTDNA.
Слева – все 64 гаплотипа гаплогруппы R1a, которые образовали плотную ветвь, составленную из субклада R1a-L657. То, что ветвь вполне однородная, показывает следующее дерево, уже только из гаплотипов данной ветви R1a-L657. Однородность дерева уже показывает, что все гаплотипы родственные, все происходят от одного общего предка, во всяком случае, с той точностью, которая нас в данном случае вполне устраивает. Посчитаем, когда этот предок жил. Во всех 64 гаплотипах в 37-маркерном формате – 924 мутации, что дает 924/64/0.09 = 160 → 190 условных поколений, то есть 4750±500 лет назад. Расчет с помощью калькулятора Килина-Клёсова, основанного совершенно на другом принципе расчета, а именно по каждому маркеру отдельно, используя калиброванные абсолютные константы скорости мутаций для каждого из 37 маркеров, дал время жизни общего предка гаплотипов R1a в Индии 4964±548 лет назад, то есть, округляя, получаем 5000±550 лет назад. Это та же величина, что и полученная «линейным методом» 4750±500 лет назад, в пределах погрешности расчетов.
Рис. 2. Дерево 64 индийских гаплотипов гаплогруппы R1a-L657 в 37-маркерном формате. Общий предок дерева жил 4750±500 лет назад. Построено по данным Индийского проекта FTDNA.
Но это время, отдаляющее нас от времени жизни общего предка гаплотипов группы R1a, живущих в настоящее время в Индии. Это вовсе не значит, что этот предок жил в Индии 4750-5000 лет назад. В Индию его потомки пришли примерно 3400-4000 лет назад, то есть на тысячу лет позже времени жизни своего общего предка. Где же этот предок жил, откуда в Индию пришли арии?
Чтобы ответить на этот вопрос, рассмотрим базовый гаплотип индийских R1a. Он – следующий (к нему, ко временам примерно 5000 лет назад, сходятся все 64 указанные гаплотипа):
25 16 10 11 14 12 12 10 13 11 17 – 16 9 10 11 11 24 14 20 32 12 15 15 16 – 11 12 19 23 15 16 18 19 35 39 13 11
А вот – предковый гаплотип этнических русских гаплогруппы R1a, общий предок которых жил на Русской равнине 4900 лет назад. Гаплотип приведен в книгах «Происхождение славян» (А.А. Клёсов, 2013, стр. 26) и «Арийские народы на просторах Евразии» (А.А. Клёсов, К.А. Пензев, 2014, стр. 246):
25 16 11 11 14 12 12 10 13 11 17 – 15 9 10 11 11 24 14 20 32 12 15 15 16 – 11 12 19 23 16 16 18 19 35 38 14 11
Мы видим, что предковые гаплотипы ариев и этнических русских очень похожи, между ними на вид всего 5 мутаций. На самом деле там всего 3.495 мутаций, потому что все отличающиеся аллели дробные. 3.495 мутаций между двумя 37-маркерными базовыми гаплотипами разделяют эти гаплотипы (то есть предков индийцев и этнических русских) на 3.495/0.09 = 39 → 41 условных поколений, или примерно 1025 лет. То есть их общий предок (субклад R1a-Z645) жил примерно за 500 лет до появления общих предков сегодняшних этнических русских гаплогруппы R1a и индийцев той же гаплогруппы, или примерно 5500 лет назад.
Примерно ту же датировку дают снипы (то есть SNP, необратимые мутации в Y-хромосоме). По данным Владимира Таганкина, который серьезно занимается снипами Y-хромосомы, между Z645 и Z283 всего два снипа, между Z283 и Z282 всего один снип, и между Z282 и Z280 – всего три снипа. «Всего» – потому что по предварительным данным один снип в среднем образуется каждые 100-150 лет назад, данные еще недостаточно откалиброваны. Поэтому при шести снипах между Z645 и Z280 (последний образовался примерно 5000 лет назад), Z645 (общий предок ариев и этнических русских) образовался примерно 5600-5900 лет назад. Они действительно близкие родственники, в пределах нескольких столетий. Более того, при наличии древней арийской топонимики на Русском севере становится ясным, что арии вовсе не замыкались в южных степях, как полагают некоторые археологи на основе материальных признаков степной и лесостепной полосы юга России и Украины. Ясно, что арии заселяли всю Русскую равнину с юга до севера, имели практически такие же гаплотипы, как и предки современных этнических русских (с общим предком всего за несколько веков до тех времен). Поэтому совершенно разумно считать носителей гаплогруппы R1a-Z645 ариями, как и их потомков, носителей субкладов Z93-Z94-L657 и Z283-Z282-Z280. Они все современники друг друга.
Теперь посмотрим, как считают популяционные генетики. Поскольку во всех 64 37-маркерных гаплотипах (то есть на 2368 маркеров) индийских носителей гаплогруппы R1a имеется 924 мутации, то они получают, используя «скорость Животовского», 924/2368/0.00069 = 566 поколений по 25 лет на каждое, то есть 14150 лет назад. Иначе говоря, при типичных расчетах попгенетиков общий предок индийских гаплотипов группы R1a жил более 14 тысяч лет назад. Какие тогда арии? Таких ариев не бывает. Поэтому попгенетики делают следующий вывод – ариев в природе не было, а гаплогруппа R1a зародилась в Индии. Как видите, применять фантастически искаженную «популяционную скорость мутаций» вовсе не безобидно. Это тянет за собой безумные «исторические интерпретации», не имеющие ничего общего с реальностью. Более того, этому рады индийские националисты, для которых «ариев придумали англичане, чтобы порабощать индийский народ». Я понимаю, что в России эта мышиная возня националистов в Индии вообще никого не интересует, да и меня тоже, но считать нужно правильно, хотя бы во имя науки. Попгенетика и здесь показала себя лженаукой.
Взглянем еще раз на «мою» страничку в Википедии. Читаем в разделе «критика»:
Носители этой гаплогруппы (R1a) в ДНК (Y-хромосома), как считает автор (А.А. Клёсов) в противовес мнению иных эксперто, перешли в Индию с севера во II тысячелетии до н.э. под самоназванием «арии»…
Итак, автор (то есть я) так «считает» (да не только автор, а тот же Гордон Чайлдс еще в 1926 году, см. выше, о чем «критики» предпочли не упоминать). Славно, не так ли? И далее:
Результаты ряда проведённых в 2000-е годы генетических исследований показали несостоятельность гипотез о доисторических миграциях индоевропейцев в Индию. См., например, следующие публикации:
Sanghamitra S., Zhivotovsky L.A., King R., Mehdi S.Q., Edmonds C.A., Chow C.-E.T., Lin A.A., Mitashree M., Sil S.K., Ramesh A., Usha Rani M.V., Thakur C.M., Cavalli-Sforza L.L., Majumder P.P., Underhill P.A. Polarity and Temporality of High-Resolution Y-Chromosome Distributions in India Identify Both Indigenous and Exogenous Expansions and Reveal Minor Genetic Influence of Central Asian Pastoralists (англ.) // American Journal of Human Genetics (англ.) русск. — The American Society of Human Genetics, 2006. — В. 78. — Т. 2. — С. 202-221.
Sanghamitra S., Singh A., Himabindu G., Banerjee J., Sitalaximi T., Gaikwad S., Trivedi R., Endicott P., Kivisild T., Metspalu M., Villems R., Kashyap V.K. A prehistory of Indian Y chromosomes: Evaluating demic diffusion scenarios (англ.) // Proceedings of the National Academy of Sciences of the United States of America. — 2006. — В. 103. — Т. 4. — С. 843-848.
Swarkar S., Rai E., Prithviraj S., Mamata J., Shweta S., Katayoon D., Bhat A.K., Bhanwer A.J.S., Tiwari P.K., Bamezai R.N.K. The Indian origin of paternal haplogroup R1a1* substantiates the autochthonous origin of Brahmins and the caste system (англ.) // Journal of Human Genetics (англ.) русск. — 2009. — В. 54. — С. 47-55. — DOI:10.1038/jhg.2008.2
Underhill P.A., Myres N.M., Rootsi S., Metspalu M., Zhivotovsky L.A., King R.J., Lin A.A., Chow C.-E.T, Semino O., Battaglia V., Kutuev I., Järve M., Chaubey G., Ayub Q., Mohyuddin A, Mehdi S.Q., Sanghamitra S., Rogaev E.I., Khusnutdinova E.K., Pshenichnov A., Balanovsky O., Balanovska E., Jeran N., Dubravka Havas A., Baldovic M., Herrera R.J., Thangaraj K., Singh V., Singh L., Majumder P., Rudan P., Primorac D., Villems R., Kivisild T. Separating the post-Glacial coancestry of European and Asian Y chromosomes within haplogroup R1a (англ.) // European Journal of Human Genetics (англ.) русск. — Macmillan Publishers, 2009. — В. 18. — С. 479-484. — DOI:10.1038/ejhg.2009.194
Metspalu M., Gallego Romero I., Yunusbayev B., Chaubey G., Mallick C.B., Hudjashov G., Nelis M., Mägi R., Metspalu E., Remm M., Pitchappan R., Singh L., Thangaraj K., Villems R., Kivisild T. Shared and Unique Components of Human Population Structure and Genome-Wide Signals of Positive Selection in South Asia (англ.) // American Journal of Human Genetics (англ.) русск. — The American Society of Human Genetics, 9 December 2011. — В. 6. — Т. 89. — С. 731-744. — DOI:10.1016/j.ajhg.2011.11.010
Я понимаю, Википедию наука не интересует, им нужно собрать все «до кучи». Но давайте разберемся. Первая статья в списке, Sanghamitra S et al. (2006), те же одиозные фамилии – Животовский, Underhill и другие. Какой метод расчета применялся? Естественно, Животовского. Какой возраст гаплотипов R1a там получили? Естественно, «exceed 10,000–15,000 years», то есть «превышающий 10-15 тысяч лет». Вывод? Никаких индоевропейцев в Индии не было. Историки и лингвисты в шоке, но не Л.А. Животовский и П.А. Андерхилл, а также под раздачу попавший бедняга Кавалли-Сфорца, член всех академий мира. Подставили его почем зря.
Следующая статья (тоже 2006) – не лучше. Группа индийцев и пара эстонцев-попгенетиков, не смыслящих в ДНК-генеалогии – Kivisild и Willems, президент эстонской академии наук, соавтор всех публикаций. Их у него, наверное, уже тысячи. Расчетов там вообще никаких нет, но есть «основополагающий» принцип – авторы полагают, что если бы арии пришли в Индию, то непременно принесли бы с собой букет разных гаплогрупп – C3, DE, J*, I, G, N, and O, а раз таких в Индии нет, то никакие арии в Индию не приходили. То, что они арии могли быть сами по себе, не смешиваться с чужаками, авторам в голову не пришло. А ведь именно это было на протяжении тысячелетий в высших кастах Индии – никаких смешиваний. Те, кто смотрели в СССР индийские фильмы в 1950-1970 гг., лицезрели индийские драмы и трагедии, как общество предохраняло высшие касты в Индии от смешивания с «низкими» кастами, и никакая межкастовая любовь не была оправданием. Так и продержались более трех тысяч лет, и сейчас гаплогруппа R1a в высших кастах достигает 72% от всех (Sharma et al, 2009). И вот эта националистическая индийская чепуха идет на «мою» страничку в Википедию как якобы критика результатов и выводов ДНК-генеалогии.
Хорошо, переходим к третьей статье (2009). Там в Википедии почти у всех авторов перепутали имена и фамилии, и Shweta Singh, например, в Википедии идет как Shweta S., как аналогично и все остальные. Ну да ладно, каков поп, в смысле, «критики», таков и приход. Что более важно, в статье опять расчеты проведены «по Животовскому». Результат – гаплогруппа R1a в Индии образовалась 13768.12 лет назад и (другой вариант) 18478.26 лет назад, если считать 25 лет на поколение, и 17623.19 и 23652.17 лет назад, если считать 32 года на поколение. Представляете, до сотой части года! Это насколько нужно быть безголовым, чтобы время до общего предка считать до сотой части года. Настоящая популяционная генетика… Здесь «метод Животовского» дает завышение в датировке в 2.75 раз и (другой вариант) в 3.70 раз. Про 32 года на поколение и говорить не приходится, это дает итоговое завышения в датировках в 3.52 и 4.73 раз. Результат – опять обсуждения того, что гаплогруппа R1a образовалась в Индии. А датировка на самом деле ниже, чем в Европе. В качестве дополнительного «аргумента» выставляется то, что «частота» гаплогруппы R1a в высших кастах Индии достигает 72%, а в европейских странах – 40%, как полагают авторы статьи. Авторы не понимают, что частота к происхождению никакого отношения не имеет. В Ирландии, например, частота гаплогруппы R1b около 92%, а гаплогруппа появилась в Южной Сибири.
Предпоследняя статья, Underhill et al. (2010, в Википедии опять ошибка), очередная «мусорная» работа в отношении расчетов, в которой изучали субклад R1a-M458. Датировка его по данным ДНК-генеалогии – примерно 4000 лет назад, образовался на границе Польши и Белоруссии. Это – западнославянский и центрально-европейский субклад, имеет две соответствующие ветви, каждой примерно по 2900 лет. Ясно, что ко времени его образования арии давно ушли из Европы на Русскую равнину, и оттуда, как носители субклада R1a-L657 – в Индию. Понятно, что эти M458 c ариями в Индию никак не могли попасть, как современные англичане никак не могли попасть в Америку с Колумбом. Но у попгенетиков свои правила, которые кроме как псевдонаукой опять же не назвать. Применяя те самые печально известные «константы Животовского», они «нашли», что возраст R1a-M458 в Польше уходит аж в мезолит, на 10700±4100 лет назад.
Эта расчетная ошибка попгенетиков во главе с Андерхиллом потащила за собой другие ошибки – они решили, что раз субклад М458 такой древний, из мезолита, то почему его нет в Индии (и в Средней Азии), куда он должен был попасть вместе с ариями? Отсюда последовал вывод, что ариев вовсе не было. То есть расчетная ошибка, бич попгенетиков, повела за собой грубую историческую ошибку.
На самом деле задачка решается в один ход – субклад М458 намного моложе субклада Z93, многочисленные потомки которого и выявлены в Индии. Практически все индийские R1a относятся к субкладу R1a-L657, возраст которого примерно 5000 лет назад (см. выше). Именно тогда в Европе только образовался субклад М458. Иначе говоря, когда R1a-L567 пришли в Индию, М458 только зарождались в Европе. Ясно, что они не могли в то же время прийти в Индию, арии из Европы давно ушли, на 500-1000 лет ранее.
К сожалению, вот таким мусором, как расчеты и исторические интерпретации в статьях попгенетиков, где есть «датировки» и соответствующие «интерпретации», заполнены академические журналы, как и цитируемый Eur. J. Human Genetics. Заметьте, там в авторах и российские попгенетики Балановские. Теперь для многих понятно, почему они «пошли свиньей» на ДНК-генеалогию, требуя ее отовсюду снять как «лженауку». Да потому что становится очевидным, что их статьи придется отзывать как ошибочные, а их, статей, – десятки, и все практически мусор в том, что касается датировок и исторических интерпретаций.
Обратите внимание, что у статьи – 34 автора, и ни один не задался вопросом, что здесь что-то не так. Вот в чем фундаментальная проблема с попгенетиками – некомпетентность и пассивность. И вот таким мусором заполнены академические издания в области попгенетики. В цитируемой статье такая обойма авторов: Underhill, Myres, Руутси, Metspalu, Животовский, King, Lin, Chow, Semino, Battaglia, Кутуев, Ярве, Chaubey, Ayub, Mohyuddin, Mehdi, Sengupta, Рогаев, Пшеничнов, Балановский, Балановская, Jeran, Augustin, Baldovic, Herrera, Thangaraj V. Singh, L. Singh, Majumder, Rudan, Primorac, Виллемс, Кивисилд. Представляете, 34 автора (российских и эстонских я выделил кириллицей, страна должна знать своих героев), и никто не понимает, что они делают, как и зачем так считают. В итоге полностью искаженное представление истории. Это же, заметим, вынесено в виде критики на «мою» страничку в Википедии, в которой делается вывод, что я неправ, и арии в Индию не приходили, и что это якобы современные представления. И дается галерея таких вот «мусорных» ссылок попгенетиков. Стыд и позор тем, кто такую ерунду написал, а редакция Википедии приняла.
Кстати, в пленарном докладе д-ра Боринской (Ин-т общей генетики РАН) на Конференции по карачаево-балкарцам, в котором половина времени (и материала) было посвящено автору этих строк, даже с показом моего портрета, она особое внимание уделила моим высказываниям из очерка «Суета попгенетиков вокруг прародителей европейцев», где я критиковал недавнюю статью («Ancient human genomes suggest three ancestral populations for present-day Europeans»), под которой подписались 98 авторов (Балановские среди них). Она в докладе представило дело так, что я якобы критиковал число авторов, мотивируя тем, что в других статьях и по 150 авторов бывает. При этом она умолчала, что на самом деле моя ключевая фраза была «я сделал критический разбор этой статьи, у которой 98 авторов, а дитя, как говорится, без глазу…». Моя критика была о том, что 98 авторов породили запутанное, вязкое исследование, изобилующее искажениями, неверными положениями, что они не состыковали геномные данные с данными ДНК-генеалогии, уже давно известными, да и просто с филогенией Y-хромосомы, которая не должна противоречить положениям и выводам статьи. Вот почему «98 авторов, а дитя без глазу». Была бы статья толковой, пусть там хоть тысяча авторов. Вот и выше – 34 автора (Underhill et al. 2010), а статья привела к неверным выводам и заключениям, которые уже несколько лет цитируют в десятках публикаций и на форумных дискуссиях, не задумываясь, что 34 автора породили мусор.
Так и со статьей с 98 авторами. Одна из выступающих с пленарным докладом на той же конференции«Этногенез, история, язык и культура карачаево-балкарского народа» (ноябрь 2014 г., РАН) объявила, что статья замечательная и продвигает наше знание в отношении происхождения европейцев. И дальше было сообщено, в чем именно продвигает – в том, что было найдено, что «происхождение древних европейцев сходится к трем древним популяциям – древние охотники-собиратели, древние фермеры, и древние евразийцы». Представляете, какой основной вывод «эпохальной» статьи 98-ми популяционных генетиков? Давайте расшифруем это замечательное высказывание. По принятым в попгенетике понятиям, «древние охотники-собиратели» – это все те, кто жили более 7 тысяч лет назад. «Древние фермеры» – это те, кто жили менее 7 тысяч лет назад. Ну, а древние евразийцы – все остальные. Переводим на нормальный язык, без этого волапюка. Вот перевод: «древние европейцы происходят от древних людей». Замечательно, не так ли? Продвигает знание. Для этого вывода понадобились 98 популяционных генетиков.
И дальше я пишу в своем очерке на Переформате:
Итак, очередная статья по анализу генома попгенетиками. Это уже означает как данность – будет очередной цирк. Но эта статья – знаковая. В ней почти сто авторов. Просто так в статьях по сто авторов не бывает. Это значит, статья заявлена как эпохальная. Еще бы – найдены предки жителей современной Европы. Три предковые популяции. И об этом ста авторами объявлено на весь мир.
А откуда сто авторов-то? Так тоже не бывает. Так раньше никогда не было. Вы видели у Эйнштейна, Менделеева, Паскаля по сто авторов в статье? Да и вообще – как сто авторов могут писать статью?
А они ее и не писали. Как поясняет сноска к статье, ее писали три автора, а по сути – только один, Иосиф Лазаридис, сотрудник Гарварда и МТИ. Еще двое, как начальники, статью просмотрели, возможно, сделали замечания – это David Reich из Гарварда и Johannes Krause из Германии. Некоторые писали отдельные части Приложения к статье. Остальные соавторы, типа все тех же Балановских, присылали образцы ДНК. Это полезно, сомнений нет, только за такую техническую работу раньше в соавторы не вставляли, а выносили благодарность в сноске. Это, конечно, мелочь, но показывает стиль современной «науки».
Переходим к заключительной, пятой статье на «моей» страничке в Википедии, из серии призванных показать, что ариев в Индии не было, и что ДНК-генеалогия поэтому показывает неверные выводы, во всяком случае в отношении Индии. Из предыдущих четырех статей мы видим, какова цена этим статьям. В трех из них применен «метод Животовского» (и в двух из этих статей в авторах сам Л. Животовский), одна – просто откровенное недоразумение, где авторы рассуждают «по понятиям, а не по науке». И вот – последняя статья. Она содержит ссылки на все четыре предыдущие статьи, разобранные выше. На них статья базируется, а именно, что ариев в Индии не было, в Индию они не приходили. То есть направление статьи уже задано. Так и оказалось. Но в этой статье уже геномные данные. И что же они показывают?
Данных много, проанализировали геномы 142 человек из 30 индийских этнических популяций, а всего расматривали геномы 1310 человек из 112 популяций. Главные выводы – что Индия в целом, по популяциям, очень древний субконтинент, с высоким «геномным разнообразием». Что выявлены два основных «предковых компонента», которые характеризуются «разнообразием и частотой» – один простирается от южной и западной Азии до Кавказа, второй, который характеризуется более высоким «разнообразием», сдвинут к южной Азии, и соответствует по численности более чем 50% индийских популяций. Его древность в целом больше, чем 3500 лет, когда арии должны были прибыть в Индию. По мнению авторов, это не согласуется с «арийским вторжением» 3500 лет назад, а также ранее или позже. Правда, постоянно оговаривается, что данные по «потоку генов» сложны и неточны, что в древности таких «потоков» было много и они смазывают картину, что согласно некоторым авторам (те, что даны в Википедии и разобраны мной выше) индийцы во множестве ушли в Европу, и что даже «поток генов» между Индией и соседней Средней Азией является сложным и трудным для анализа.
Заключение статьи звучит так (убирая пустые в данном контексте фразы, например, о том, что процесс генетического структурирования в южной Азии продолжается и сейчас): «Получены сложные данные, которые трудно объяснить только предполагаемым недавним прибытием индо-ариев, но который свидетельствует о многоообразных потоках генов в южно-азиатский генетический пул, как с запада, так и с востока, на протяжении долгого времени». И дальше – про липидный метаболизм и диабет, и что нужно продолжать заниматься исследованиями. Ну и где там данные, противоречащие прибытию ариев в Индию?
Напоминаю, под каким «соусом» это помещено на «моей» страничке в Википедии. А именно, что это – «в противовес мнению иных экспертов», в смысле в противовес данным ДНК-генеалогии, и что это показывает «несостоятельность гипотез о доисторических миграциях индоевропейцев в Индию». Отается только развести руками. Пять статей «иных экспертов», три из которых откровенно неверны, так как применяли псевдонаучные «популяционные скорости», завышающие датировки на сотни процентов, одна статья полна недоразумений и построена «по понятиям», последняя вообще ничему не противоречит. Ох уж эта Википедия, которая в угоду «рецензентам», не имеющим никакой квалификации, засоряет ерундой в общем-то ценное издание.
Несколько комментариев в отношении последней статьи, по геному индийцев. На самом деле то, то авторы получили, никакого удивления не вызывает. Я уже не раз в своих статьях показывал, что, как ни странно, картина снипов в геноме следует за простенькой Y-хромосомой. Действительно, в геноме определяются сотни тысяч и миллионы снипов, а в ДНК-генеалогии Y-хромосомы для каждой гаплогруппы – всего несколько, определяющие гаплогруппу и субклад. То есть как бы хвост вертит собакой, причем хвостик совсем маленький. Любой генетик скажет – что за ерунда, мы Y-хромосому вообще почти не принимаем во внимание, когда анализируем геном. Вот в чем опять драма и трагедия современной генетики вообще и попгенетики в частности. Они не смотрят «трехмерно», за пределы своей маленькой (в этом отношении) парадигмы. А там четкие закономерности, которые генетики не видят, потому что не знают и не хотят знать филогению гаплогрупп Y-хромосомы. Связь между геномом и филогенией Y-хромосомы я описал на ряде примеров в статье «Действительно ли «генетики нашли разных русских?». Статью я закончил так: «…(надо) понять, что лежит в основе вариаций в геноме, задающих глубинные различия между популяциями. Эти различия идут из тьмы тысячелетий, и что особенно интригует – они определенно завязаны на мутации, определяющие мужские гаплогруппы. Это – совершенно новая концепция, и решать эту загадку надо в содружестве генетиков и ДНК-генеалогов».
Так вот, то, что в Индии «древнее разнообразие» – совершенно не удивительно, его диктуют, в частности (и в особенности) древние гаплогруппы L, H, J, Q, C, R2, и каждую из них можно датировать с помощью подходов ДНК-генеалогии. То, что в этом «разнообразии» тонет относительно недавняя гаплогруппа R1a в Индии – неудивительно, хотя ее около трети в современной Индии, но это только среди тех, кто может заплатить не менее 200 долларов за тест. Индийцы из джунглей в это число не входят. Поэтому вряд ли число носителей R1a в Индии превышает 15% от всего мужского населения. Это и есть в основном потомки ариев, прибывших в Индию примерно 3500 лет назад. А те, кто образует геномную связь с Афганистаном и Кавказом – это носители гаплогрупп G2a, L1b и L1c, наряду с другими, минорными. В общем, все эти геномные данные можно усилить данными ДНК-генеалогии, в том числе корректными датировками, и привести их в порядок.
Похоже, что в Индии осталось небольшое количество особенно древних носителей гаплогруппы R1a, и они иногда выявляются в джунглях, но систематических исследований их не проводилось, субклады их не определялись, в высших кастах (и в кастах вообще) они не представлены, они попадают при классификации или в низшие касты (lower caste), или в племена (tribes). Видимо, они попали в Индию в ходе древнейших миграций из Южной Сибири на запад, когда носители R1a в итоге прибыли в Европу примерно 9-10 тысяч лет назад (Klyosov, 2009-2012). Но эти данные получены на коротких гаплотипах, в которых ошибки тестирования проявляются намного больше, чем при работе с протяженными гаплотипами. А поскольку данные по этим древним R1a в Индии фрагментарные (в известном списке Sengupta [Sengupta et al., 2006] их, например, практически нет, за исключением, быть может, гаплотипов 4, 95, 109 на дереве гаплотипов на рис. 3), то систематического изучения их пока не проводилось, субклады неизвестны.
Рис. 3. Дерево из 110 10-маркерных гаплотипов гаплогруппы R1a в индийских кастах и племенах. Построено по данным Sengupta et al. (Amer. J. Human Genetics, 2006).
Формат гаплотипов – DYS 393, 390, 19, 391, 388, 439, 389-1, 393, 389-2, 461).
На данных Сенгупты (2006) стоит остановиться чуть подробнее, поскольку именно «расчеты» этих самых известных данных по Индии (более тысячи 10-маркерных гаплотипов в выборке, из них 110 гаплотипов гаплогруппы R1a) приводят популяционных генетиков к тому, что общий предок носителей R1a в Индии жил более 14 тысяч лет назад (Животовский – в авторах и статьи Sengupta et al; естественно, датировки опять рассчитывали «по Животовскому»). Дерево гаплотипов на рис. 3 не слишком симметричное, но считать вполне можно, на что указывает и полученный базовый гаплотип дерева (здесь приведены 9 маркеров, поскольку десятый, DYS461, не входит в список 37-маркерных гаплотипов, которые будут приведены ниже для сравнения, Х – маркеры, которые не определяли в работе Sengupta)
25 15 10 Х Х Х 12 10 13 11 17
Все 110 гаплотипов содержат 344 мутации, что дает 344/110/0.018 = 174 → 211 условных поколений, то есть 5275±600 лет до общего предка. Здесь 0.018 – константа скорости мутации для 9-маркерного гаплотипа, равная сумме констант для всех 9 маркеров, определенных еще в 2006 году Чандлером (Chandler, J. Genet. Geneal., 2006): 0.00076 + 0.00311 + 0.00151 + 0.00265 + 0.00022 + 0.00477 + 0.00186 + 0.00052 + 0.00242 = 0.01782 мутаций на гаплотип на поколение.
Сравним полученный базовый гаплотип с таковым для индийцев субклада R1a-L657, приведенным выше в 37-маркерном формате (общий предок жил 4750±500 лет назад):
25 16 10 11 14 12 12 10 13 11 17 – 16 9 10 11 11 24 14 20 32 12 15 15 16 – 11 12 19 23 15 16 18 19 35 39 13 11
Если переписать его в виде, удобном для сравнения
25 16 10 Х Х Х 12 10 13 11 18
то мы увидим, что базовый гаплотип, полученный по данным Сенгупты (включающих низшие и высшие касты и племена), и базовый гаплотип, полученный по данным индийского проекта FTDNA, различаются всего на одну мутацию (выделено), а на самом деле всего на 0.55 мутаций; если усреднить по всем аллелям маркера DYS19, получим DYS19 = 15.55. Другими словами, это одинаковые предковые гаплотипы, и одинаковая датировка, в пределах погрешности расчетов, безотносительно, 9-маркерные гаплотипы или 37-маркерные. Некоторое завышение датировки по данным Сенгупты, которое, впрочем, укладывается в погрешности, вызвано тремя особенно мутированными гаплотипами (под номерами 4, 95, 109, см. рис.3), на которые приходится 19 мутаций, и при их снятии получаем 325/107/0.018 = 169 → 203 условных поколений, или 5075±580 лет до общего предка.
Такова цена утверждениям попгенетиков, что ариев в Индии не было, и что общий предок индийских R1a жил более 14 тысяч лет назад. И здесь ДНК-генеалогия полностью переигрывает популяционную генетику.
которая усугубляется фальшивыми «калибровками» (примеры)
В первой части этой статьи был дан иллюстративный пример расчета времени общего предка знаменитой шотландской «генеалогической» семьи МакДоналдов, которая в гаплогруппе R1a ведет свою линию от Джона Лорда Островов (John Lord of the Isles), умершего в 1386 году, то есть 628 лет назад. Было показано, что подход ДНК-генеалогии дает совершенно разумное совпадение с документальной генеалогией по Джону и его потомкам.
Документальная генеалогия дает 628 лет до годовщины смерти Джона (1386 год), а ДНК-генеалогия дает 650-700 лет до времени его жизни (1314-1364 год), то есть практически абсолютное совпадение в контексте данного исследования, учитывая, что год его рождения неизвестен.
Попробуем рассчитать те же данные «по Животовскому»:
6-маркерные гаплотипы: 17/68/6/0.00069 = 60 поколений, то есть 1500 лет до Джона.
12-маркерные гаплотипы: 44/84/12/0.00069 = 63 поколения, то есть 1575 лет до Джона.
25-маркерные гаплотипы: 109/84/25/0.00069 = 75 поколений, то есть 1875 лет до Джона.
37-маркерные гаплотипы: 178/59/37/0.00069 = 118 поколений, то есть 2950 лет до Джона.
Столь разные величины поколений и лет в результате применения «метода Животовского» получаются потому, что константы скорости мутаций для разных гаплотипов разные (самые высокие – для 37-маркерных гаплотипов), а в «методе Животовского» они для всех одинаковые – 0.00069 мутаций на маркер на 25 лет. Но это не самая большая проблема (хотя она приводит к дополнительным ошибкам почти в два раза, от 1500 лет до 2950 лет). Еще хуже то, что результаты получаются «ни в какие ворота». Не говорите об этом расчете генеалогам семейства МакДоналдов, засмеют и выгонят, откажутся разговаривать. Вот так работают «расчеты» популяционных генетиков. Как видим, разница та же, завышение в датировке на 230-450%, в данном конкретном случае. А поскольку случаи разные, то попгенетики задирают датировки обычно в 2.3-5.0 раз. Вот такая цена их расчетам.
«Знающий» попгенетик возразит – не может быть, Животовский свои скорости мутаций калибровал, это все популяционные генетики знают, и основополагающая статья его 2004 года основана на калибровках его скорости мутации, той самой, 0.00069 мутаций на 25 лет на маркер. Там всё доказано, потому его константу и взяли за основу в популяционной генетике.
И вот здесь начинается самое интересное. Похоже, ни один популяционный генетик, ни один (!), не удосужился проверить сам, лично, что там была за «калибровка». Ни один не взял исходные данные Животовского и не проверил. Я это сделал. Давайте посмотрим, как Л. Животовский калибровал. Основная его «калибровка» – по группе болгарских цыганских таборов, 8-маркерные гаплотипы; по группе полинезийцев, 10-маркерные гаплотипы; и по группе африканских банту. Идея такая – мы знаем, что цыгане появились в Европе примерно 900-1000 лет назад, в Болгарии – 700 лет назад, поэтому если взять группу их гаплотипов, посчитать мутации, приложить «популяционную» скорость 0.00069 мутаций на маркер на 25 лет, и если получится примерно 700 лет для Болгарии, или 900-1000 лет для Европы (Хорватии, например), то скорость 0.00069 – правильная. Разумно, не так ли?
Именно это Л. Животовский и объявил, что при калибровке так и получилось, именно со скоростью мутации 0.00069, так и вошло в учебники. То же самое в отношении полинезийцев и банту – у Животовского тоже получилось как надо, скорость 0.00069. А как же, калибровка-то вот она, налицо, так все уже 10 лет и повторяют. Так и ответили авторы статьи (Hammer, Животовский и др., 2009) на мои развернутые комментарии в журнале Human Genetics (Klyosov, 2009), в которых я показал, что они считают неверно, ошибаются в 3-4 раза. Авторы сослались на «the original Zhivotovsky et al. (2004) study (i.e., the effective rate was calibrated for time periods of *1,000 years). Since 2004, this approach has been employed by other researchers and critically evaluated in the scientific literature». Перевод: «Исходное исследование Животовского и др. (2004) (содержало) калибровку получающихся скоростей на протяжении примерно 1000 лет». Типа, какие могут быть вопросы, и тем более критика? Скорости калибровались, и всё тут.
Проверим, чего, похоже, никто не делал. Такая «наука» у популяционных генетиков. Давайте посмотрим для начала на цыган. Гаплотипы цыган были определены у 179 человек из двенадцати болгарских таборов. На удивление, у всех гаплотипы похожи, явно от общего не столь далекого предка. Посмотрим сначала, как надо работать с этими гаплотипами, а потом сверим, как «калибровал» Животовский. Для двойной проверки проанализируем как 6-, так и 8-маркерные гаплотипы, и если считаем правильно, результаты должны совпасть, в пределах разумных погрешностей.
У всех 179 цыган гаплотипы в 6-маркерном формате содержали 34 мутации от базового гаплотипа 15 12 22 10 11 12 (в формате DYS 19, 388, 390, 391, 392, 393). Это дает 34/179/0.0088 = 22±4 поколения до общего предка, то есть 550±100 лет. Поскольку из 179 гаплотипов 146 были идентичными друг другу, то есть базовыми, то логарифмический метод дает [ln(179/146)/0.0088] = 23±3 поколения до общего предка, то есть 575±75 лет назад. Практически полное совпадение результатов по линейному и логарифмическому методу. Это означает, что, действительно, все 179 цыган (или их подавляющее большинство) произошли от одного общего предка.
В 8-маркерном формате (дополнительно вошли DYS 389-1 и 389-2) добавилось 25 мутаций, то есть стало 59 мутаций. Получаем: 59/179/0.013 = 25±4 поколения до общего предка, то есть 625±100 лет. В 8-маркерном формате из 179 гаплотипов 126 базовые. Получаем [ln(179/126)/0.013] = 27±4 поколения до общего предка, то есть 675±100 лет назад.
Как видим, все четыре варианта расчета дали расстояние до общего предка болгарских цыган одно и то же, в пределах погрешностей расчета: 550±100, 575±75, 625±100 и 675±100 лет назад. Надо отметить, что хотя присутствие цыган в Болгарии было отмечено 700 лет назад, но эти сведения не обязательно точны, они могут относиться к другим цыганам, не обязательно гаплогруппы Н1 (как в данном случае), потомство у первых цыган могло не выжить, и так далее. Тем не менее, 550-675 лет назад – это не слишком далеко от отмеченного времени «начала присутствия» цыган в Болгарии.
Читатель, видимо, уже приготовился, что у Животовского получилось значительно больше, то есть не менее 1500 лет до общего предка цыган, и что он повинился, что такого науке не известно, значит, ошибочка вышла с константой 0.00069. Слишком занижена. Ведь цыган было 179, мутаций (в 8-маркерных гаплотипах) 59, так что у Животовского должно получиться 59/179/8/0.00069 = 60 поколений, то есть 1500 лет до общего предка. Но наиболее понимающие читатели, зная популяционных генетиков, уже заулыбались. Потому что у тех раз результат заранее известен, такой и получится. Наука такая. Надо просто уметь рукава пониже держать, желательно, чтобы руки закрывали.
Следим за рукавами. Один гаплотип был из списка удален, потому что он «оказался нечетным» (?). Какой удалять – естественно, на усмотрение удаляющих (или базовый, или мутированный, то есть уже направленно вносится искажение серии данных). Далее постулировали, что общий предок всех цыган жил 700 лет, то есть 700/25 = 28 поколений назад. Замечаете прием? Я-то выше рассчитывал, когда жил общий предок (получилось в среднем 600 лет назад), а Л. Животовский просто постулировал, что 700 лет назад, и отсюда вел отсчет. Но есть большая разница – время, когда цыгане были замечены в Болгарии, и время, когда жил общий предок рассматриваемых 179 цыган. Совсем не обязательно, что он был первым цыганом в Болгарии, как уже отмечено выше.
Но 600 и 700 лет – разница не так велика, так как же он подтвердил свою скорость мутации в три раза ниже?! А вот как – после совершенно произвольного снятия одного гаплотипа получилось, что при использовании квадратичного метода среднее число мутаций на маркер было 0.01272, и при делении на постулированные 28 поколений до общего предка средняя скорость мутации на маркер оказалась 0.01272/28 = 0.000454. А надо 0.00069, непорядок. Надо данные менять, иначе «теория» под угрозой. Животовский с коллегами написали в статье, что это была «недооценка». Поэтому решили пересчитать данные, введя некие «веса», и в ходе этого сняли еще ряд гаплотипов цыган. После каждой стадии подгонки смотрели, насколько скорость удалось подогнать к 0.00069, но никак не получалось. Тогда решили, что один табор – «Музыканты» – «очень гетерогенный» (на самом деле, по моим расчетам, он был совершенно нормальный, как показано выше при сопоставлении линейных и логарифмических методов), и сняли его целиком, все 19 гаплотипов, и опять ввели «веса по размеру популяции» (что ни в коем случае нельзя делать). Получили среднюю скорость мутации 0.000725±0.000187 (обратим внимание на величину погрешности), и решили, что это и есть 0.00069.
Если это «калибровка», то непонятно, что такое калибровка в понимании авторов «основополагающей» статьи, на которую все попгенетики обязаны ссылаться, иначе рецензенты статью в журнал не пропускают. На это жаловался венгерский автор на форуме по «генетической генеалогии», о чем я уже писал на Переформате. Вот такая популяционная генетика. На самом деле, авторы зря занимались акробатикой. Раз надо получить 0.00069, то способ простой – надо было повыкидывать гаплотипы с мутациями как «гетерогенные» или просто дефектные, или те, которые просто не понравились, оставить на оставшиеся полторы сотни человек ровно 23 мутации, и получить 23/150/8/0.00069 = 28 желаемых поколений, то есть как раз искомые 700 лет. «Калибровка» была бы не хуже, и никаких хлопот. Как говорил хрестоматийный герой – «как пожелаем, так и сделаем».
Так же рассчитывали и по гаплотипам полинезийцев (маори, самоанцы и жители островов Кука). Решили исходить из того, что маори прибыли в Новую Зеландию 800 лет назад, то есть 32 поколения назад, по 25 лет на поколение. Гаплотипы были 10-маркерные, но акробатика пошла и там. Самоанцев решили в расчет не включать, а у остальных сократить гаплотипы до 7-маркерных. Видимо, с 10-маркерными не получалось то, что «пожелали». Применяя квадратичный метод, нашли, что среднее число мутаций на маркер в 7-маркерных гаплотипах у маори и островитян Кука равно 0.00998, и разделив на 32 поколения, получили таким образом «форсированную» (в отличие от естественной) скорость мутации 0.00032. Как видно, это вовсе не 0.00069, а в два с лишним раза медленнее. Авторы заключили, что это опять «недооценка», и снова пустились в долгие и вязкие рассуждения, почему это может быть недооценкой. На этом «калибровка» данной серии гаплотипов была завершена.
С африканскими банту и того хуже. Раздел «Материалы и методы». Смотрим про африканских Банту. Читаем – «анализировали 148 человек гаплогруппы Е3а7-М191», перечисляются племена, сообщается, что анализ проводили по 10-маркерным гаплотипам.
Раздел «Результаты». Данных по Банту нет, они вообще в данном разделе не упоминаются.
Раздел «Обсуждение». Банту не упоминаются.
Раздел «Применение». Говорится, что применили полученную величину 0.00069 к популяции Банту, чтобы посмотреть, как это коррелирует с археологическими, лингвистическими и историческими данными. Далее говорится, что если усреднить по всем 11 племенам и популяциям, перечисленным в разделе «Материалы и методы», то получится 3400±1100 лет до «экспансии» Банту в подгруппе со снипом М191. И далее – «археологические и лингвистические данные поддерживают это значение, поскольку показывают присутствие Банту в Западной Африке в неолитические времена ∼1000 лет до нашей эры, или даже 2000 лет до нашей эры». И далее – «Величина 3400 лет назад может рассматриваться как нижняя граница для времен экспансии Банту… Если мы возьмем наиболее частые гаплотипы среди тех 148 хромосом как предковые, и приложим к ним величину 0.00069, то время для М191 получится 14700 лет… В этом случае, экспансия Банту могла произойти ранее, чем 3500 лет назад». И далее идут долгие рассуждения с неопределенной концовкой. Остается совершенно загадочным, причем здесь 14700 лет назад, когда усреднение дало 3400 лет назад «до экспансии», и причем здесь вообще «экспансия». Иначе говоря, авторы просто перешли на другую тему, и про «калибровку» успешно забыли. Нет её там.
Если это «калибровка», то что такое не калибровка? К сожалению, исходные данные (гаплотипы) в статье не приведены, ссылка статьи, что они «в сети», не дает никакого линка. Я могу только заключить, что рассматриваемая популяция Банту молодая, не более 1000-1200 лет до общего предка, как это часто бывает с африканскими популяциями, только недавно прошедшими очередное бутылочное горлышко. Ни к каким археологическим или лингвистическим данным это не имеет никакого отношения. Рассуждения в статье не имеют никакого значения в отношении рассматриваемого вопроса. Никакой «калибровки» не было и близко.
Такова цена «калибровкам» Л.А. Животовского, которые стали краеугольным камнем популяционной генетики в последние 10 лет. Кроме как «стыд и позор» других слов я не подберу. Последние пять лет я объясняю это в научной литературе, начиная со статьи в Journal of Genetic Genealogy (2009) и Human Genetics (2009), и далее в журнале Advances in Anthropology, выступаю на англоязычных форумах. Но попгенетикам – как о стену горох. Они продолжают плодить в академической литературе мусор. А те, кто в этом ровным счетом ничего не понимают, как тот же воронежский «индолог» Семененко, тем не менее, вылезают со своим критическим словом, что «метод Животовского» самый правильный, потому что им пользуются все популяционные генетики в мире. И далее – «Я опираюсь на данные международных коллективов профессиональных генетиков, опубликованные в ведущих международных научных журналах по генетике». Грош цена этим «международным коллективам профессиональных генетиков», а также рецензентам этих «международных научных журналов», а также неучам, кто это бездумно повторяет.
Напомним, что Л. Животовский применил совершенно усредненные и искусственные приближения, приняв, что константа скорости мутаций в Y-хромосомах ДНК должна быть усреднена по 10 тысячам (!) гаплогрупп за все время существования человечества, и тогда у него получилось, что она, константа скорости мутации, должна быть равна 0.00069 мутациям за 25 лет на маркер, причем она одинакова для любых гаплотипов, 6-, 7-, 8-, 9-, 10-, 12-маркерных и так далее. Любой, кто имеет хотя бы минимальный опыт работы с гаплотипами знает, что для каждого маркера есть своя константа скорости мутации, и в 6-маркерные гаплотипы попадают одни маркеры, в 8- или 9-маркерные свои (и, значит, средняя константа скорости мутации будет уже другой), в 10- или 12-маркерные гаплотипы добавляются еще другие маркеры со своими константами, и так далее. Считать, что константа одна на все – это, во-первых, принципиально неверно, во-вторых – это вносить значительную погрешность в расчеты. Именно эту ошибку делают все без исключения «международные коллективы профессиональных генетиков», чьи фундаментально ошибочные статьи «опубликованы в ведущих международных научных журналах по генетике».
Это – трагедия современной популяционной генетики. А люди, которые в этом совершенно не разбираются, продолжают кудахтать, другого слова не подберу, что «он не генетик» (это про меня). Но генетика здесь совершенно не при чем, нет в ДНК-генеалогии генетики, генетика закончилась до ДНК-генеалогии, когда генетики определили, какие мутации есть в ДНК, сколько «тандемных повторов» есть в маркерах, и какие снипы найдены в каких участках Y-хромосомы. Иначе говоря, генетика, а точнее, лаборанты в компаниях типа FTDNA или в лаборатории популяционной генетики РАМН, снабжают нас этими данными, а ДНК-генеалогия идет дальше, проводя количественную обработку этих данных, чего генетики делать уже не умеют. У них образование не то, их этому не учили. А учиться, как показывают последние 6-8 лет, они либо не могут, либо не хотят.
Что же касается того, что наши с И.Л. Рожанским калибровки констант скоростей мутаций «не дали ничего нового», и «просто повторили давно известное» (то, что заявил Балановский с трибуны в ходе своего доклада), то это только указывает на уровень компетенции «специалиста». Уровень Балановского – ужасающе низкий. Совершенно серый уровень, никакой творческой искры, сплошные штампы.
Кстати, у меня на конференции был продолжительный разговор с Л.А. Животовским, в ходе которого был снят целый ряд недоразумений, как-то застрявших в голове Льва Анатольевича. Он выставил мне четыре вопроса (они-то и были недоразумениями), и остался ответами полностью удовлетворен, вопросы снял. Один из них отражал традиционное заблуждение (скорее, дезинформацию) попгенетиков, что я якобы приравниваю гаплогруппу к этносу. Как только я напомнил, что везде пишу, что у этнических русских есть четыре основных гаплогруппы и полтора десятка минорных, у карачаево-балкарцев три основных гаплогруппы и целый ряд минорных, он тут же сказал – понял, вопрос снимаю.
Другой вопрос был о том, почему я не учитываю «пропадания» генеалогических линий, поскольку по причине этого он и использует сильно заниженную константу скорости мутации (0.00069, а не 0.00179 для 67-маркерных, или 0.00243 для 37-маркерных), чтобы компенсировать потери. И еще, почему я не учитываю размер популяции, поскольку когда у отца рождается в итоге 40 сыновей, а у тех еще по много сыновей, то идет быстрый разбег мутаций в ДНК сыновей и прочих внуков и правнуков, и это надо учитывать. Об этом он рассказывал в своем докладе.
Я пояснил, что это принципиальное заблуждение популяционных генетиков, потому что когда, например, между общим предком и мной 40 поколений, то мне все равно, сколько у меня братьев и прочих родственников, поскольку мутации между общим предком и мной идут исключительно на протяжении этих 40 поколений, только по этой вертикали, по этой генеалогической ниточке. У остальных выживших родственников от того же общего предка – то же самое, те же 40 поколений, как и у всех остальных. Поэтому у всех нас сегодня, у каждого потомка этого общего предка, в 111-маркерных гаплотипах в среднем по 8 мутаций. У одних 7 мутаций, у других 9 мутаций, но в среднем 8 мутаций у каждого. Если нас, протестированных, собрать 100 потомков от этого общего предка, то на всех придется 800 мутаций в наших 111-маркерных гаплотипах. Вот и получаем: 800/100/0.198 = 40 поколений до общего предка (я здесь для простоты пренебрегаю возвратными мутациями). А у тысячи потомков будет 8000 мутаций, и получим 8000/1000/0.198 = 40 поколений, то же самое. А у 20 потомков будет 160 мутаций, и получим 160/20/0.198 = 40 поколений, то же самое. И это не зависит, нас только 20 человек выжило в потомках, или 100, или 1000 человек (просто погрешность расчетов больше у малого числа гаплотипов). Потому что мутации в каждом из нас индивидуальны. Здесь, кстати, 0.198 – это константа скорости мутации в 111-маркерных гаплотипах.
Лев Анатольевич долго мучился, пытаясь побороть в себе это старое заблуждение попгенетиков, и я, чтобы ему помочь, привел пример с радиоактивным распадом. Там неважно, образец радиоактивного материала в килограмм весом, или в грамм, или в миллиграмм. Время полураспада будет одинаковым, и, значит, и константа скорости распада будет той же самой. Процесс-то первого порядка, как и мутации в гаплотипах. По такому же принципу размер образца (число потомков) не влияет на константу скорости мутации, и у попгенетиков – принципиальное заблуждение на этот счет. В итоге Л.А. согласился, но сказал, что еще помоделирует, что я приветствовал. Мы даже условно договорились написать совместную статью, в которой разобрать и сравнить основные принципы и положения популяционной генетики и ДНК-генеалогии, но позже Л.А. Животовский от такой статьи отказался. Нетрудно догадаться, почему.
Правда, несколько позже выяснилось и другое. В первой части этой статьи я уже писал, что на Конференции была зачитана (профессором Э.К. Хуснутдиновой) Резолюция Круглого стола по антропологии, генетике и ДНК-генеалогии, в которой не было никаких слов «осуждения» ДНК-генеалогии, да их и не могло быть, поскольку аудитория принимала ДНК-генеалогию с энтузиазмом, и даже изгнала из зала Балановских, которые пытались устроить обструкцию дополнительной лекции по ДНК-генеалогии, на которую собрался зал. Иначе говоря, был провал противников ДНК-генеалогии.
Однако Балановская изготовила подметную «резолюцию», и разместила ее еще 27 ноября на сайте своей лаборатории популяционной генетики РАМН, то есть именно в день своего изгнания, когда для Конференции оставался еще полный день работы. Далее она начала проталкивать этот подметный вариант, который на Конференции не звучал, через Оргкомитет Конференции и докладчиков. Концовка его следующая: «в рамках распространяемой в интернете «ДНК-генеалогии» отождествляются принципиально разные этнические и генетические категории, прикрепляются этнические названия к генетическим «родам» и археологическим культурам. Понятийно-расчётный аппарат ДНК-генеалогии научно необоснован, не прошёл научную апробацию в индексируемых научных журналах и основан на смешении популяционно-генетических, генеалогических и историко-культурных понятий. Все эти черты ДНК-генеалогии выводят ее за рамки академических наук и находятся в противоречии с необходимостью корректного подхода, различающего биологические данные о популяции от понятия народа во всей его сложности. Поэтому упоминание ДНК-генеалогии в названии Круглого стола неуместно».
Спустя 10 дней, Балановская прислала эту «резолюцию» мне для «согласования», тем самым фактически подтвердив, что на ее сайте была размещена фальшивка, а не согласованная Резолюция. При этом она сообщила, что «Проект резолюции разработан учеными, представляющими и генетику, и антропологию: Е.В. Балановской, О.П. Балановским, Л.А. Животовским, Д.В. Пежемским. Также текст был одобрен В.А. Шнирельманом, мнение которого особенно важно в плане формулировок, касающихся этносов. Окончательные формулировки были утверждены председательствовавшим модератором (Н.К. Янковским)».
Характерно, что фамилий профессора Э.К. Хуснутдиновой и профессора М.Д. Каракетова, основных модераторов Круглого стола, среди перечисленных «разработчиков» нет. Пикантно и то, что именно Э.К. Хуснутдинова зачитывала настоящую Резолюцию на заключительном пленарном заседании Конференции, но не попала в список «разработчиков». Как и М.Д. Каракетов, который пытался остановить Е. Балановскую в ходе ее доклада, когда она давала мне персональные (негативные, разумеется) характеристики, но остановить ему не удалось. Балановская его откровенно проигнорировала, и, как видно, к составу «разработчиков резолюции» не подпустила. Зато там оказался В.А. Шнирельман, которого не было ни среди модераторов, ни среди докладчиков Круглого стола, и который, уверяю, не имеет никакого понятия о ДНК-генеалогии.
Кстате о В. Шнирельмане. Совершенно очевидно, что его привлекли к «одобрению» не случайно. Он – автор печально известного доклада при Московском бюро по правам человека, доклада под названием «Мифы современного расизма в РФ», в котором он как пугало выставляет древних ариев, раздел доклада называется «Праиндоевропейцы и русский национализм», и весь доклад построен в издевательских конструкциях в отношении изучения древней истории русского народа. «Расисты» для Шнирельмана – это те, кто занимается древней историей славян, их он называет «идеологами», и они, конечно, Шнирельману неприятны. Он пишет:
…они объявляют, что «вопрос древней истории – это первостепенный вопрос государственной стратегии». Со своей стороны они рисуют Евразию исконной родиной славяно-русов, где они жили якобы в течение тысячелетий, закладывая здание будущей человеческой цивилизации. Подхватывая мессианский советский миф, эта концепция опрокидывает его в отдаленное прошлое и изображает доисторических предков богатым и щедрым народом, отважными воинами и неутомимыми завоевателями, благородными культуртрегерами и создателями едва ли не всех древнейших цивилизаций.
Разумеется, это Шнирельману крайне неприятно. В список таких «идеологов», осуждаемых Шнирельманом, попали известный востоковед И.М. Дьяконов и лингвисты В.В. Иванов и Т.В. Гамкрелидзе, а также «мифотворец-патриот Л.Н. Гумилев» (о, патриот – это бранное слово у Шнирельмана), сибирский археолог В.Е. Ларичев («пропагандировались фантазии сибирского археолога В.Е. Ларичева о древнейшей в мире цивилизации в Сибири, созданной, естественно, индоевропейцами»), писатель В.А. Чивилихин (который «настаивал на том, что предки славян были автохтонами в поволжских и причерноморских степях. В итоге он договаривался до того, что славяне будто бы существовали как общность уже пять тысяч лет назад», и «не без удовольствия замечал, что и в долине Хуанхэ древнейшее население было представлено светлокожими индоевропейцами» (какое уж там удовольствие для Шнирельмана…).
В число «идеологов» попали также академик Б.А. Рыбаков, в трудах которого «рисовалась многотысячелетняя история первобытных славян» (это – крайне неприятно для Шнирельмана), как и то, что «корни славян Рыбаков издавна искал в бронзовом веке», и «Рыбаков делал все, что в его силах, чтобы обнаружить славянские корни в глубинах первобытности». Шнирельману крайне неприятно то, что академик Рыбаков «полагал, что доказательство глубокой древности славян положительно скажется на самосознании и самоощущении русского народа. Не случайно одну из своих популярных статей он назвал «глубокие корни – могучая крона»». С точки зрения Шнирельмана, это плохо. Это – плохая «идеология». Это – «национализм». Интересно, с каких пор шнирельманы диктуют нам, русским, что такое «национализм», и почему русский патриотизм – это плохо, это плохая «идеология».
В число «идеологов» попал и О.Н. Трубачев, который, по мнению Шнирельмана, является «важным источником информации для русских националистов», а именно его «теория… о близком родстве и теснейших контактах между славянами и индоариями в Северном Причерноморье». И далее – «…Трубачев всеми силами пытался доказать, что после ухода оттуда основной массы их соплеменников в Переднюю Азию какие-то группы индоариев надолго задержались в Северном Причерноморье и вполне могли иметь тесные контакты с ранними славянами». Это «не осталось незамеченным в стане русских националистов». «В своих псевдонаучных построениях они …следовали разработкам Иванова и Гамкрелидзе», а другие «вновь вернулись к идее балканской прародины». По мнению Шнирельмана, это очень плохо, ну, не могут древние русские и прочие славяне быть «щедрым народом, отважными воинами и неутомимыми завоевателями», это все вредные мифы. А уж арии – это пунктик у Шнирельмана. Арии непременно относятся к нацистской Германии. И так далее, полсотни страниц вот такого ненавистного по отношению к истории славян текста. Может, это и есть расизм, только шнирельмановский?
Надо сказать, что за прошедшие со времени его доклада несколько лет основные его пассажи в отношении отсутствия древней истории славян оказались полностью отвергнуты если не «официальной наукой», то научными данными, в первую очередь ДНК-генеалогией. Подтвердились выводы и В.Е. Ларичева, и И.М. Дьякова, и Б.А. Рыбакова, и О.Н. Трубачева, и В.В. Иванова и Т.В. Гамкрелидзе, и Л.Н. Гумилева, и В.А. Чивилихина, но никак не Шнирельмана.
Понятно, что он с готовностью принял приглашение Балановской, для того, чтобы «текст был одобрен В.А. Шнирельманом, мнение которого особенно важно». Ну, понятно, не бином Ньютона. Для Балановской и Шнирельмана древность славян – как нож по сердцу. Поэтому надо «валить и мочить» ДНК-генеалогию. Вот так делаются дела в РАН, и наглость тех, кто так «делает», не имеет предела. История опять повторяется, в виде фарса.
Так вот, я написал Л. Животовскому, и спросил, действительно ли он был среди «разработчиков» этой «резолюции». Он ответил, что проект резолюции ему прислали в начале декабря (то есть когда этот «проект» уже неделю подавался Балановскими в СМИ как якобы принятый Конференцией, то есть окончательный), и по сообщению Л. Животовского, он, то есть Животовский, вычеркнул из «проекта» слова «выводят ее (т.е. ДНК-генеалогию) за рамки академических наук», а все остальное оставил. То есть оставил и «в рамках распространяемой в интернете «ДНК-генеалогии»», и «прикрепляются этнические названия к генетическим «родам» и археологическим культурам», и «понятийно-расчётный аппарат ДНК-генеалогии научно необоснован», и «не прошёл научную апробацию» и «основан на смешении популяционно-генетических, генеалогических и историко-культурных понятий», и «находятся в противоречии с необходимостью корректного подхода».
И это после того, как он снял практически все свои вопросы в ходе нашей с ним дружеской дискуссии в очереди в академической столовой и за самим обедом. Все вопросы снял! А если не снял – имел все возможности снять. Он же знает прекрасно про мои статьи и книги в области ДНК-генеалогии, но поддерживает слова «распространяемой в интернете»! Он прекрасно знает, что я не прикрепляю никаких «этнических названий» к археологическим культурам, да и как это возможно? У них уже есть названия. Скажем, майкопская культура – как можно к ней применять «этническое название»? – эти названия все молодые, а культура – старая. Как Л. Животовский может поддерживать «положение» о том, что расчетный аппарат ДНК-генеалогии «научно необоснован»? Что он «находится в противоречии с необходимостью корректного подхода»? Все это Животовский прекрасно знает, но… Помните, что Ленин говорил про интеллигенцию, что это вовсе не мозг нации? Вот то-то и оно. Это он и про Животовского говорил. На меня, признаться, Л. Животовский при личном общении поначалу произвел хорошее впечатление. Видимо, по сравнению с Балановской, которая произвела при личном общении впечатление крайне отталкивающее. Но, увы, с Животовским произошла ошибка. Манеры забили сущность. Нерукопожатность не выявилась. Поначалу.
Возвращаемся к «методу Животовского» и «генеалогическому методу» в исполнении попгенетиков. Первый – вообще чудовищно неправилен в любом исполнении, второй в исполнении попгенетиков просто примитивен, и потому неправилен. Там не делят серии гаплотипов на ветви, там смешивают общих предков ДНК-линий, там получают фантомные датировки, в общем, все равно получается ерунда. Но поскольку Балановские думать не умеют, то они в последнее время на всякий случай используют оба метода, так в статьях и описывают – «метод Животовского», и «генеалогический метод». А потом подбирают, какой им лучше подойдет, по обстановке. Это что, наука? Позор это, а не наука. Разобраться они так и не смогли, и, естественно, их датировки в таком бессильном варианте никому не нужны. Разница их – обычно в три раза друг с другом. Вот и судите об уровне их компетенции и «творческой» пассивности.
В 2009 году в журнале Human Genetics вышла статья группы авторов – Michael Hammer, Doron Behar, Tatiana Karafet, Fernando Mendez, Brian Hallmark, Tamar Erez, Lev Zhivotovsky, Saharon Rosset, Karl Skorecki – по датировке общего предка коэнов (по Библии и Торе – потомков Аарона, брата Моисея, которого (Аарона) Всевышний назначил быть первосвященником. Эта работа была мной задолго до того выполнена, и я показал, что общий предок коэнов (гаплогруппы J1) жил чуть больше тысячи лет назад (1,075±130 лет назад – линейным методом, и примерно 1025 лет назад – логарифмическим). Понятно, что по датировке нынешние коэны этой гаплогруппы никакого отношения к Аарону не имеют, поскольку библейский Аарон должен был жить примерно 3600 лет назад, во времена библейского же исхода из Египта. Мои данные показали, что, возможно, их предки прошли «бутылочное горлышко популяции» и продолжили свою ДНК-линию в 10-м веке нашей эры. Но, по предположению Президента международной ассоциации коэнов в нашей с ним личной переписке, возможно, тот уважаемый еврей тысячу лет назад был просто назначен коэном, или избран местной еврейской общиной, поскольку общине был нужен легитимный лидер, а наследственного коэна, чтобы продолжить дело, не оказалось. Такая процедура предусмотрена иудейским религиозным законодательством. Поэтому общий предок нынешних коэнов и жил примерно тысячу лет назад, в 10-м веке нашей эры плюс-минус сотня лет.
Но указанная группа авторов (Hammer, Животовский и др.) применили, разумеется, «скорость Животовского», и получили, разумеется, «время жизни Аарона» в библейские времена, а именно 3190±1090 лет назад, о чем торжественно заявили, что получили прекрасное совпадение с ожидаемым результатом. Статья закачивается так: «Полученная датировка согласуется с гипотезой, что «линия коэнов» является уникальной ДНК-линией древних евреев, и унаследуется по отцовской линии вместе с линией еврейского духовенства». Там же обязательные фразы, что история еврейских священников сложна, и ее необходимо продолжать изучать.
Поскольку результаты и выводы статьи были фундаментально ошибочными, я написал откровенно разгромную статью в тот же журнал, Human Genetics, на 14 страницах, в которой подверг резкой критике «метод Животовского» и полученные результаты, и пересчитал заново все их данные, приведя совершенно красноречивые деревья гаплотипов. Статья была принята к печати через неделю после ее получения редакцией. Ответ попгенетиков ждали больше месяца, задержав выпуск журнала. Ответ пришел совершенно беззубый, вялый и неконструктивный. И понятно почему – крыть авторам было нечем, можно было только вяло возражать, не приводя никаких данных или разумных аргументов. «Аргументы» были следующими, в сумме их было восемь:
1. Оппонент (то есть я) выступил с критикой слишком быстро, что «не есть в интересах основательного научного процесса».
2. В свой критике я ссылаюсь на недоступные и неопубликованные работы (в печати в J. Genetic Genealogy).
3. Использую нестандартную терминологию, как, например, «логарифмический» и «линейный» методы.
4. Я сделал неверный вывод, что гаплотипы образуют «генеалогию» и к ним могут применяться «генеалогические» подходы (то есть не «популяционные»).
5. Я не учитывал «генетический дрейф» или «никакие другие особенности мутаций в микросателлитах».
6. Авторы не знают, насколько мой метод применим к их системе.
7. Применяемый метод Животовского правильный, откалиброван и критически рассмотрен в научной печати.
8. Применяемые мной «деревья гаплотипов» являются интересными и могут быть в высшей степени полезными, но к ним нужно относиться с «большой осторожностью».
Положение (1), конечно, вызывает смех. Авторы, видимо, не в курсе, что «метод Животовского» уже давно вызывает насмешки, что я его давно и детально разобрал (что и делаю опять в данной статье), и на это описание не нужно времени.
Положение (2) можно было бы принять, если бы не русскоязычные Л. Животовский и Т. Карафет в авторах. Один вопрос мне в частной переписке, если они не умеют пользоваться Интернетом, и я бы дал им ссылки на работы, на которые они легко могли бы взглянуть. Мог бы и выслать. Если бы они хотели. Но желания, явно, не было.
Положение (3) – авторы просто не владеют предметом. Логарифмический метод в химической кинетике существует уже десятки лет. Он подробно и в азбучном варианте изложен в учебнике И.В. Березина и А.А. Клёсова «Практический курс химической и ферментативной кинетики» (1976), который уже почти 40 лет служит основным учебником по данному предмету для университетов. Опять – короткий вопрос со стороны Л. Животовского или Т. Карафет, и они узнали бы для себя много нового. Но не захотели.
Положение (4) – авторы опять не поняли, о чем речь, или сделали вывод, что не поняли. А речь о том, что дерево гаплотипов можно разложить по ветвям, и к каждой ветви приложить «генеалогический подход» и «генеалогическую скорость мутаций». А вот обрабатывать все дерево чохом, поперек ветвей, как делают авторы, категорически неверно.
Положение (5) – авторы снова не поняли, что разложение дерева по ветвям – это и есть учет «генетического дрейфа». Фраза «Никакие другие особенности мутаций в микросателлитах» может только вызвать смех. Какие «другие особенности» – авторы не пояснили.
Положение (6) – в моем «Комментарии» всё описано. Авторы просто не захотели, или не смогли разобраться.
Положение (7) – неверно и подтасовочно. Мы уже знаем, какой он «правильный», как «откалиброван» и как «критически рассмотрен в научной печати».
Положение (8) – наконец-то, разумные слова. Следует, правда отметить, что к «методу Животовского» слова про «осторожность», по мнению авторов, видимо, неприменимы. Там осторожность не нужна. Это только к другим авторам относится. Во всяком случае, прошло более пяти лет, и слова «являются интересными и могут быть в высшей степени полезными» так и остались словами. Судя по сегодняшней реакции Балановских, эти слова они к действию не приняли. Напротив, ДНК-генеалогия у них вызывает озлобление, и я уже пояснил, почему. Если в 2009 году они еще были начинающими, то сейчас погрязли в неверных данных, статьях и интерпретациях, пройдя точку невозврата.
С тех пор я многократно критиковал «метод Животовского» в печати – и в Вестнике Академии ДНК-генеалогии, и в J. Genet. Geneal., и в книге «Происхождение славян», в главе 16, и в журнале Advances in Anthropology. Этот «метод» неоднократно критиковал Dienekes Pontikos в своем блоге, подчеркивая его фундаментальную неприменимость для расчетов по сериям гаплотипов. Но попгенетики литературу не читают и литературы не знают, кроме своих собственных статей.
со стороны попгенетики в научной печати
Этот раздел исключительно краткий. Просто потому, что в научной печати никакой критики ДНК-генеалогии не было. Вся «критика» сводится к совершенно неквалифицированным, а зачастую злобным комментариям в сети (Балановские, Дробышевский, Семененко и многие другие «любители»). То, что на ноябрьской Конференции в РАН в своих докладах говорили Боринская и Балановские, это не критика, это набор в лучшем случае недоразумений и искажений, в худшем – откровенная ложь. Об этом – в следующих частях этой статьи.
Продолжение следует…
Анатолий А. Клёсов,
доктор химических наук, профессор
Перейти к авторской колонке
| Продолжается сбор средств на проекты ДНК-генеалогии! Ваша поддержка – это дальнейшее изучение настоящей истории наших предков, выпуск тематических книг, организация научных мероприятий, исследование палео-днк и ещё многое другое. Ключевой проект – издание учебника А.А. Клёсова «ДНК-генеалогия. Практический курс». Давайте вместе продвигать новое научное знание –кнопки для оплаты пожертвования внизу по этой ссылке, буквально один клик. Можно перевести пожертвование от 100 до 5000 руб. любым удобным для вас способом. Также сообщаем о новой книге А.А.Клёсова и К.А. Пензева, которую можно приобрести в книжных магазинах или онлайн. |