Саморекламы нынче много, но есть самореклама умеренная, а есть
помпезная, нелепая, вызывающая смех. Например, вот эта, спекулятивная,
неквалифицированная, примитивная попытка решения важных вопросов истории
под громкими лозунгами «решения эпохальных проблем». Конечно,
эпохальные проблемы могут быть решены в рамках исследования ДНК, но для
этого подходы должны быть адекватными поставленным задачам, а не быть
скороспелыми и надуманными, и фактически дублировать то, что уже
известно, более того, подгонять к тому, что уже известно.

Например, О. Балановский, работающий в области популяционной генетики, на днях провозгласил, что после статьи его и коллектива из 39 человек, включающих обоих (или обеих?) Балановских, Касьяна, Дыбо и других, плюс 39 человек из Генографического консорциума, под названием «Генетическое наследство популяций, говорящих на балто-славянских языках: синтез аутосомных, митохондриальных и Y-хромосомных данных» (Kushniarevich et al., сентябрь 2015) – «в ближайшее десятилетие вряд ли можно ожидать чего-то принципиально нового в области генетики и лингвистики славянских популяций». Рассмотрим коротко, так ли это.
Начнем с того, что с самого начала статьи грубо нарушается принцип, наиболее последовательно сформулированный академиком В.В. Седовым – «Совместные решения этногенетических проблем представителями разных наук возможны только при том условии, что выводы каждой отрасли науки покоятся на собственных материалах, а не навеяны данными смежной науки». В статье же в самом начале приводится дерево балто-славянских языков (см. ниже), и вся статья ставит своей задачей подгонку генетических данных под это дерево. Да, собственно, подгонка и не понадобилась, потому что подгонять было нечего, поэтому дело ограничилось отдельным обсуждением лингвистики, и отдельным – генетики, оба в крайне примитивном исполнении.
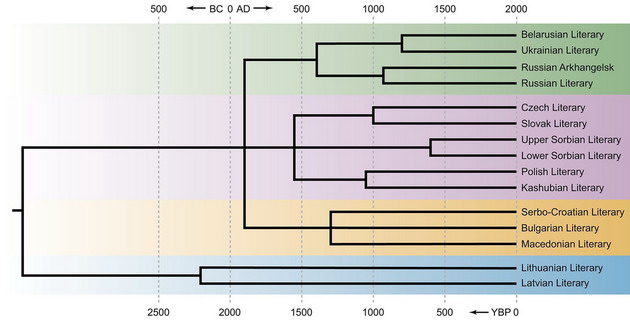
Дерево балто-славянских языков (из статьи Kushniarevich et al., 2015)
Если бы авторы были знакомы с ДНК-генеалогией, то им бы не понадобилось громоздкое «генетическое исследование», в котором компьютер анализировал массивы данных по аутосомам (то есть «поперек всей ДНК»), митохондриальным ДНК и Y-хромосоме. В итоге компьютер прокручивал информацию и выдавал некие формализованные показатели, наносил на график, и этот график шел в статью. Так работают популяционные генетики, обычно не думая, что все это означает. Остается только написать, что «полученные данные подтверждают дерево балто-славянских языков».
Взглянем на гаплогруппный состав в славянских и балтских популяциях. Современные славяне сложились в основном из трех родов, или гаплогрупп – R1a, I2a и N1c1. Остановимся кратко на том, что в ДНК-генеалогии называется родом, этот разумный и корректный термин приводит Балановского в исступление. Гаплогруппа – это совокупность родственных субкладов, происходящих от еще более древних общих предков, а субклад – это совокупность носителей одних и тех же снипов. Раз снипы одни и те же, то ясно, что все их носители, то есть в данном случае мужчины, произошли от одного общего предка, патриарха, в ДНК которого этот снип впервые образовался. Фактически это есть общепринятое определение рода, что есть совокупность всех поколений людей, происходящих от одного предка.
Ясно, что при определении рода речь может идти только о роде или по мужской линии, или по женской, поскольку смешанных родов быть не может. Вспомним генеалогические деревья предков, они обычно строились по мужской линии, потому что введение женской линии делает дерево уже при нескольких поколениях вниз практически нечитаемым. Соответственно, ДНК-генеалогия рассматривает род или по мужской линии, то есть по Y-хромосоме, или по женской, по митохондриальной ДНК. Здесь мы будем рассматривать только рода по мужской линии. В терминах ДНК-генеалогии эти основные рода, или рода верхнего уровня, называются гаплогруппами, которые, в свою очередь, подразделяются на множество субкладов. Ряд субкладов охватывают сотни миллионов человек, и часто тоже называются гаплогруппами, как, например, R1a и R1b, хотя, строго говоря, их гаплогруппой (верхнего уровня) является гаплогруппа R. Поэтому четкого разделения между понятиями «гаплогруппа» и «субклад» нет, использование того или иного термина определяется контекстом.
Как и у субкладов, в основе каждой гаплогруппы был общий предок, патриах гаплогруппы, в Y-хромосоме которого произошла определенная мутация, присущая только данной гаплогруппе. Естественно, эта мутация присутствует у носителя каждого субклада в данной гаплогруппе. Например, гаплогруппа R1a, доминирующая гаплогруппа (род) этнических русских, включает 66 субкладов (по классификации ISOGG, международного общества, которое занимается регистрацией гаплогрупп и субкладов, и регулярно вносит изменения и дополнения в классификацию), а гаплогруппа R1b, доминирующая в Западной и Центральной Европе, насчитывает 471 субкладов. Обе гаплогруппы произошли примерно в одно и то же время, но вторая гаплогруппа изучается значительно бóльшими научными силами, потому и субкладов идентифицировали большее количество. Обе гаплогруппы входят в гаплогруппу R1, а та – в гаплогруппу R, гаплогруппу верхнего уровня.
В итоге гаплогруппы (верхнего уровня) не просто соответствуют определённым родам, но образуют определённую последовательность, лестницу гаплогрупп, показывающих их иерархию. Об этом мы много раз говорили, не будем повторяться.
Вернемся к славянам и балтам – а именно, в нашем случае, к литовцам и латышам, и продемонстрируем их родовой состав с точки зрения ДНК-генеалогии. Относительно минорные гаплогруппы показывать не будем, чтобы не загромождать приводимые данные, и поясним это на следующем примере.
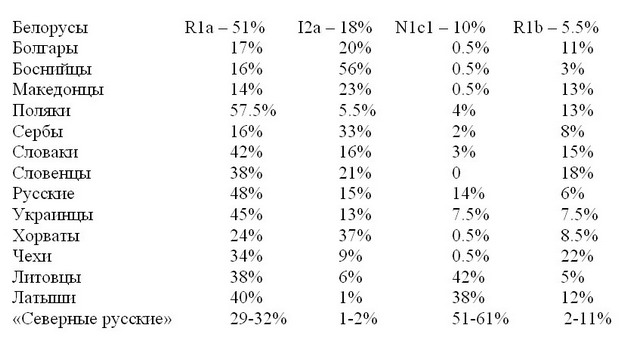
Основной состав белорусов по гаплогруппам следующий:
R1a – 51%
I2a – 17.5%
N1c1 – 10%
У белорусов еще есть 5.5% I1, 5.5% R1b, 4% E1b, 2.5% J2, 1.5% G, 1% J1, 1% I2b, но ни одна из них не является характерно славянской. I1 – древнеевропейская гаплогруппа, или гаплогруппа Старой Европы (до III тыс. до н.э.), почти исчезнувшая в III тыс. до н.э., прошедшая «бутылочное горлышко популяции» и возродившаяся во II тыс. до н.э., и ставшая общеевропейской, как было описано выше; R1b – наследие ямной культуры и отчасти сарматов, потомков того же древнего рода R1b, прошедшего миграциями от Сибири до Днепра и далее через Кавказ в Месопотамию и затем в Европу разными миграционными путями, основной из них через Пиренеи и далее в континентальную Европу как культура колоколовидных кубков; E1b – потомки древних египтян и затем древних греков, расселившиеся по Средиземноморью и черноморским берегам; J2 – потомки древних бедуинов, составившие затем в заметной части население Средиземноморья и значительную часть евреев и арабов; J1 – преобладающая гаплогруппа у евреев и арабов, а также потомков древних евреев по всему миру, включая, например, горских евреев на Кавказе и значительную часть евреев-ашкенази, передвинувшихся в средние века (в особенности, после эпидемии черной чумы середины XIV века) во все славянские страны и нашедших там убежища от преследований; G – древнеевропейская гаплогруппа, еще одна из основных гаплогрупп Старой Европы, практически исчезнувшая, остатки которой бежали в Малую Азию, на Кавказ и в Переднюю Азию, и сейчас одна из основных кавказских гаплогрупп (например, до 75% у осетин); I1b – минорная гаплогруппа по всему миру, в Европе нигде не превышает 4-7% (в основном, в Северной Европе – Бельгия, Нидерланды, Дания, Норвегия, Швеция, северная Франция, северная и западная Германия), возможно, тоже реликт Старой Европы.
Примерно такие же вкрапления указанных минорых гаплогрупп явно неславянского происхождения есть по всем странам, включая славянские. Итак, перейдем к славянским странам, добавив балтов – литовцев и латышей, а также «северных русских», как, например, коми и марийцы, которые славянами, строго говоря, не являются, но которых популяционные генетики часто используют в качестве «этнических русских», и более того, заявляют их как эталон этнических русских, чтобы поднять в последних долю «финно-угорской компоненты». Именно в таком виде российские популяционные генетики Е. и О. Балановские заявили «геномный эталон этнических русских» в международную геномную базу данных, подробнее см. здесь. Так русские оказались в базе данных «финно-уграми».
Помимо этого, добавим в список гаплогруппу R1b, для славян нехарактерную, и которая унаследована в России от ямной культуры (найдена в составе ископаемых ДНК с археологическими датировками 2700-3300 гг. до н.э., Haak et al, 2015), отчасти от намного более поздних сарматов, в Европе – от потомков культуры колоколовидных кубков (позже 4800 лет назад).
Если не уходить в детали, то картина в целом ясна – у славян преобладает гаплогруппа R1a (за исключением балканских славян – болгар, сербов, хорват, македонцев, боснийцев, у которых доминирует I2a), за ней следует характерная южнославянская гаплогруппа I2a, и замыкает южно-балтийская гаплогруппа N1c1, которая к северо-востоку становится финно-угорской. Отдельно стоят балты – литовцы и латыши, у которых содержание R1a и N1c1 практически одинаково. Как мы видим, корни славян в отношении их ДНК-родового происхождения уходят в разные стороны.
Эти данные уже позволяют полностью предсказать, что покажут громоздкие данные «генетики», полученные по миллионам точек, и обработанные крайне формализовано. Результат же заранее известен. Ясно, что поляки, белорусы, русские и украинцы, то есть «восточные и западные славяне», у которых доминирует гаплогруппа R1a, образуют один «кластер» точек, в котором русские с более заметным вкладом гаплогруппы N1c1 будут несколько смещаться в сторону балтов (литовцев и латышей), еще больше туда будут смещаться «северные русские», которые уже смыкаются с финно-уграми. Напротив, сербы, хорваты, македонцы и боснийцы, то есть «южные славяне», у которых доминирует гаплогруппа I2a, образуют отдельный «кластер», а чехи и словаки, у которых, в отличие от других, повышено содержание гаплогруппы R1b (22% и 15%, соответственно), характерной для Западной и Центральной Европы, тяготеют к немцам, с еще более высоким содержанием этой гаплогруппы, причем чехи (с более высоким содержанием R1b) тяготеют больше, чем словаки.
Именно так и получилось в статье Кушнаревич и Балановских с соавторами. Цитируем:
>> «Восточные славяне – русские из центральных и южных регионов, белорусы и украинцы – образуют свой кластер»,
>> «напротив, русские из северных областей европейской части России отходят от остальных восточных славян, и располагаются вблизи своих финно-говорящих соседей»;
>> «чехи, и в меньшей степени словаки, сдвинуты по направлению к немцам и другим западноевропейцам»;
>> «поляки или совпадают с восточными славянами, или находятся близко к ним»;
>> «большинство южных славян… балканского полуострова… отделены от остальных балто-славянских популяций, и образуют размытую группу популяций, которые разделяются на западные регионы (словенцы, хорваты и боснийцы) и восточные регионы (македонцы и болгары)».
Действительно, смотрим на состав гаплогрупп македонцев и болгар – он практически идентичен по всем четырем основным гаплогруппам, приведенным выше.
>> «латыши и литовцы, говорящие на балтских языках, расположены вблизи эстонцев, говорящих на финно-угорских языках, если рассматривать Y-хромосому, и слегка сдвинуты к восточным славянам по аутосомам».
Тоже ясно, почему, у них поровну R1a (доминирующей у восточных славян) и N1c1 (доминирующей у финно-угров). Аутосомы добавляют «женский вклад», как видно, совсем незначительный, тем более в контексте обсуждаемой работы.
>> «мордвины показывают близость к обеим балтским популяциям, видимо, отражая исторические свидетельства, что племена, говорившие на балтских языках, расширялись на восток Восточно-Европейской равнины».
Первое само собой разумеется, поскольку у мордвин в среднем заниженное значение R1a (36%), по сравнению с большинством восточных славян и почти равное таковому у литовцев и латышей (38% и 40%) при почти том же значении N1c1 (15%), что у этнических русских. А второе – вообще неверно, это не балты продвигались на восток, а, напротив, носители гаплогруппы N1c1 продвигались на запад, и на этом пути достигли Балтики, что проявляется в составе гаплогрупп литовцев и латышей.
Так что «генетика» в данном исследовании ничего нового не привнесла в то, что давно известно по спискам опубликованных распределений гаплогрупп в славянских и соседних популяциях. И какой же вывод делают авторы из того, что давно известно? Вывода три (цитаты):
>> «начальное разделение протославян осталось невыясненным»,
>> «по лексикостатистическим данным разделение на западных, восточных и южных славян произошло примерно 1900 лет назад, дальнейшая диверсификация – примерно 1300-1500 лет назад, и расхождение на отдельные языки – 1000-500 лет назад».
>> «Y-хромосомная диверсификация на низшем уровне 1 популяционной структуры – среди локальных популяций, говорящих на одних и тех же языках – варьируется от почти 0 для чехов и македонцев до 0.05 в пределах северных русских, что в среднем дает 0.01. Генетическая дифференциация среди этнических популяций, относящихся к уровню 2 лингвистической ветви, приблизительно равна 0.03, и вариация в пределах ветви уровня 3 балто-славянских популяций увеличивается до 0.06».
Всё. На этом раздел статьи заканчивается. В этом состоит, по словам О. Балановского, то, что «в ближайшее десятилетие вряд ли можно ожидать чего-то принципиально нового в области генетики и лингвистики славянских популяций».
Видимо, этот «приципиально новый вклад» состоит, как полагают авторы обсуждаемой статьи, в том, что имеет место «корреляция» между деревом языков, приведенных в начале раздела, с географией, то есть «южными славянами», «северными», «западными» и «восточными». А состав гаплогрупп тоже определяется географией, поэтому было бы более чем странно, если бы такой корреляции не было. В итоге авторы статьи используют «циклический аргумент», когда территория определяет состав гаплогрупп, она же определяет разделение языков на локальные, а значит, состав гаплогрупп определяет разделение языков на локальные. Видимо, это и есть «принципиально новое».
Вот в описанном выше и состоит главное отличие популяционной генетики от ДНК-генеалогии. И дело не только в принципиально разной методологии, дело в принципах постановки и решения задач.
На обсуждении «данных лексикостатистики» можно и не останавливаться, ничего нового там нет, обработка данных крайне формализованная и невнятная. Да и в обсуждении филолингвистического дерева, приведенного выше, у авторов нет ничего нового. Они ссылаются на дерево Gray and Atkinson (2003):
Фрагмент дерева, относящийся к балто-славянским языкам (из статьи Gray and Atkinson, 2003)
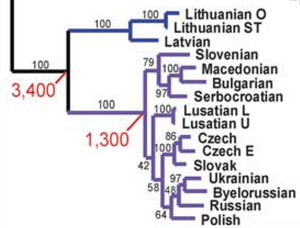
но различия (или сходства) даже не обсуждают. Что говорить, когда нечего говорить? Лексикостатистика – вообще методология, мало кем в науке принятая, из-за обилия произвольных приближений и допущений, и авторы это еще раз показывают. В любом случае, никакой новой связи с генетикой не обнаружено, а то, что описано в статье – тривиально. Но давайте коротко остановимся на показанных деревьях, на данных лексикостатистики, и данных ДНК-генеалогии по балтам и славянам. О роли лексикостатистики в исследованиях, помимо замечания выше, можно сказать, что ей, как правило, отдается весьма низкий ранг достоверности, и ее данные могут служить только как вспомогательные, как дополнительное свидетельство в пользу того, что уже получено другими методами. Как мы видим, у Грея и Аткинсона (2003) расхождение славянских и балтских языков (литовского и латышского) датируется 3400 лет назад, то есть серединой II тыс. до н.э. То же воспроизводит и дерево в статье Кушнаревич и Балановских с соавторами (2015).
Это – крайне маловероятно и не стыкуется с данными ДНК-генеалогии, согласно которым в те времена будущие балты были еще на Урале или только начинали миграции на запад от Урала. Разумеется, никаких индоевропейских языков у них тогда не было. Были уральские, угорские или те, как это решили назвать лингвисты. К балтийскому побережью предки литовцев и латышей гаплогруппы N1c1 подойдут только к середине I тыс. до н.э. Датировки по 67-маркерным гаплотипам литовцев и латышей гаплогруппы N1c1 показывают время жизни их общих предков 2600±300 и 2100±300 лет назад, соответственно. Общий предок современных этнических русских гаплогруппы N1c1 жил 3300±340 лет назад, но это включает более древние зауральские и уральские гаплотипы, поэтому и общий предок несколько древнее. В Литве и Латвии такие пока не обнаружены.
Поэтому никакого «балто-славянского единства» или «единства гаплогрупп N1c1 и R1a» в середине II тыс. до н.э., и тем более ранее, просто не могло быть, как и «славянских языков, выделившихся из балтских». Ситуация была обратной – прибывшие на берега Балтийского моря предки литовцев и латышей, видимо, относительно малочисленные носители гаплогрупп N1c1 переняли ИЕ языки автохтонных (видимо, можно так назвать) носителей гаплогруппы R1a, которые обитали там как минимум тысячу лет (а если учесть ископаемую гаплогруппу R1a на Онежском озере с датировкой 7500 лет назад, в середине VI тыс. до н.э., то обитавшие к тому времени и 5 тысяч лет). Они и образовали «единство R1a и N1c1», которое с тех пор не распадалось, поскольку и сейчас у литовцев и латышей этих гаплогрупп поровну, примерно по 40% от всех. Языки разошлись, и, как будет показано ниже из данных С.А. Старостина, русский и литовский разошлись только в первой половине нашей эры.
В связи с этим с некоторым изумлением воспринимается основное положение лингвистов, специалистов по балтским языкам, что «балто-славянское единство иногда считают продуктом позднейшего сближения двух независимых индоевропейских диалектов», ссылка. Аналогично, давно затвержденное положение об особой индоевропейской архаичности литовского языка надо понимать, что архаичным был язык носителей гаплогруппы R1a, у которых предки литовцев этот язык переняли.
Вывод – славянские языки (и их предки) не «выделились» из балтских, а, напротив, были источником ИЕ языков для предков литовцев и латышей второй половины I тыс. до н.э. – первой половины I тыс. н.э.
Если мы обратим внимание на лексикостатистику в том варианте, с которым работал С.А. Старостин, то она, на первый взгляд, дает разумные результаты. Например, согласно его данным (Старостин, 1989), в современном русском и персидском языках в стословнике Сводеша имеется 28% совпадений. Это дает
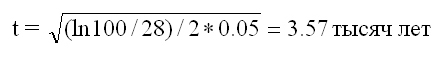
от времени расхождения языков (до настоящего времени), что примерно соответствует времени ухода ариев на Иранское плато (0.05 – это по Старостину константа скорости выпадания слов из стословника ИЕ языков). Для русского и древнеиндийского языка имеется 54% совпадений, что соответствует
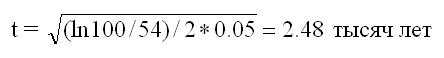
от времени расхождения языков. Это примерно равно времени, когда Панини создал классический санскрит. Какова бы ни была надежность этих интерпретаций, данные намного отличаются от степени расхождения русского и литовского языков, чему С.А. Старостин относил величину 74-76% (по разным текстам). Это соответствует времени расхождения 1660-1740 лет назад, то есть III-IV вв. нашей эры. Это никак не стыкуется с лексикостатистическими оценками ни Грея с Аткинсоном, ни Кушнаревич с сотрудниками. Не может три четверти совпадения слов из стословника соответствовать расхождению языков, начиная со столь давнего времени, как середина II тыс. до н.э.
Таким образом, мы видим, что подгонка данных «генетики» под заранее заданную, и, скорее всего, неверную датировку не только грубо нарушает принцип, сформулированный В.В. Седовым, но и ведет к неверным результатам как лингвистики, так и генетики. Основная ирония в том, что провозглашенная Балановским максима «в ближайшее десятилетие вряд ли можно ожидать чего-то принципиально нового в области генетики и лингвистики славянских популяций» оказалась опровергнута уже через две недели после этого провозглашения.
Литература:
Клёсов А.А. Действительно ли «генетики нашли разных русских»?
Седов В.В. Происхождение и ранняя история славян. М., 1979.
Старостин С.А. Сравнительно-историческое языкознание и лексикостатистика. Лингвистическая реконст-рукция и древнейшая история Востока. М., 1989. С. 3-39.
Gray R.D., Atkinson Q.D. Language-tree divergence times support the Anatolian theory of Indo-European origin. Nature, 2003. 426, 435-439.
Kushniarevich, A., Utevska, O., Chuhryaeva, M., Agdzhoyan, A., Dibirova, K., Uktveryte, I., Möls, M., Mulahasanovic, L., Pshenichnov, A., Frolova, S., Shanko, A., Metspalu1, E., Reidla, M., Tambets, K., Tamm, E., Koshel, S., Zaporozhchenko, V., Atramentova, L., Kučinskas, V., Davydenko, O., Goncharova, O., Evseeva, I., Churnosov, M., Pocheshchova, E., Yunusbayev, B., Khusnutdinova, E., Marjanović, D., Rudan, P., Rootsi, S., Yankovsky, N., Endicott, P., Kassian, A., Dybo, A., The Genographic Consortium, Tyler-Smith, C., Balanovska, E., Metspalu1, M., Kivisild, T., Villems, R., Balanovsky, O. (2015) PLOS One, September 2, DOI: 10.1371/journal.pone.0135820
Анатолий А. Клёсов,
доктор химических наук, профессор
Например, О. Балановский, работающий в области популяционной генетики, на днях провозгласил, что после статьи его и коллектива из 39 человек, включающих обоих (или обеих?) Балановских, Касьяна, Дыбо и других, плюс 39 человек из Генографического консорциума, под названием «Генетическое наследство популяций, говорящих на балто-славянских языках: синтез аутосомных, митохондриальных и Y-хромосомных данных» (Kushniarevich et al., сентябрь 2015) – «в ближайшее десятилетие вряд ли можно ожидать чего-то принципиально нового в области генетики и лингвистики славянских популяций». Рассмотрим коротко, так ли это.
Начнем с того, что с самого начала статьи грубо нарушается принцип, наиболее последовательно сформулированный академиком В.В. Седовым – «Совместные решения этногенетических проблем представителями разных наук возможны только при том условии, что выводы каждой отрасли науки покоятся на собственных материалах, а не навеяны данными смежной науки». В статье же в самом начале приводится дерево балто-славянских языков (см. ниже), и вся статья ставит своей задачей подгонку генетических данных под это дерево. Да, собственно, подгонка и не понадобилась, потому что подгонять было нечего, поэтому дело ограничилось отдельным обсуждением лингвистики, и отдельным – генетики, оба в крайне примитивном исполнении.
Если бы авторы были знакомы с ДНК-генеалогией, то им бы не понадобилось громоздкое «генетическое исследование», в котором компьютер анализировал массивы данных по аутосомам (то есть «поперек всей ДНК»), митохондриальным ДНК и Y-хромосоме. В итоге компьютер прокручивал информацию и выдавал некие формализованные показатели, наносил на график, и этот график шел в статью. Так работают популяционные генетики, обычно не думая, что все это означает. Остается только написать, что «полученные данные подтверждают дерево балто-славянских языков».
Взглянем на гаплогруппный состав в славянских и балтских популяциях. Современные славяне сложились в основном из трех родов, или гаплогрупп – R1a, I2a и N1c1. Остановимся кратко на том, что в ДНК-генеалогии называется родом, этот разумный и корректный термин приводит Балановского в исступление. Гаплогруппа – это совокупность родственных субкладов, происходящих от еще более древних общих предков, а субклад – это совокупность носителей одних и тех же снипов. Раз снипы одни и те же, то ясно, что все их носители, то есть в данном случае мужчины, произошли от одного общего предка, патриарха, в ДНК которого этот снип впервые образовался. Фактически это есть общепринятое определение рода, что есть совокупность всех поколений людей, происходящих от одного предка.
Ясно, что при определении рода речь может идти только о роде или по мужской линии, или по женской, поскольку смешанных родов быть не может. Вспомним генеалогические деревья предков, они обычно строились по мужской линии, потому что введение женской линии делает дерево уже при нескольких поколениях вниз практически нечитаемым. Соответственно, ДНК-генеалогия рассматривает род или по мужской линии, то есть по Y-хромосоме, или по женской, по митохондриальной ДНК. Здесь мы будем рассматривать только рода по мужской линии. В терминах ДНК-генеалогии эти основные рода, или рода верхнего уровня, называются гаплогруппами, которые, в свою очередь, подразделяются на множество субкладов. Ряд субкладов охватывают сотни миллионов человек, и часто тоже называются гаплогруппами, как, например, R1a и R1b, хотя, строго говоря, их гаплогруппой (верхнего уровня) является гаплогруппа R. Поэтому четкого разделения между понятиями «гаплогруппа» и «субклад» нет, использование того или иного термина определяется контекстом.
Как и у субкладов, в основе каждой гаплогруппы был общий предок, патриах гаплогруппы, в Y-хромосоме которого произошла определенная мутация, присущая только данной гаплогруппе. Естественно, эта мутация присутствует у носителя каждого субклада в данной гаплогруппе. Например, гаплогруппа R1a, доминирующая гаплогруппа (род) этнических русских, включает 66 субкладов (по классификации ISOGG, международного общества, которое занимается регистрацией гаплогрупп и субкладов, и регулярно вносит изменения и дополнения в классификацию), а гаплогруппа R1b, доминирующая в Западной и Центральной Европе, насчитывает 471 субкладов. Обе гаплогруппы произошли примерно в одно и то же время, но вторая гаплогруппа изучается значительно бóльшими научными силами, потому и субкладов идентифицировали большее количество. Обе гаплогруппы входят в гаплогруппу R1, а та – в гаплогруппу R, гаплогруппу верхнего уровня.
В итоге гаплогруппы (верхнего уровня) не просто соответствуют определённым родам, но образуют определённую последовательность, лестницу гаплогрупп, показывающих их иерархию. Об этом мы много раз говорили, не будем повторяться.
Вернемся к славянам и балтам – а именно, в нашем случае, к литовцам и латышам, и продемонстрируем их родовой состав с точки зрения ДНК-генеалогии. Относительно минорные гаплогруппы показывать не будем, чтобы не загромождать приводимые данные, и поясним это на следующем примере.
Основной состав белорусов по гаплогруппам следующий:
R1a – 51%
I2a – 17.5%
N1c1 – 10%
У белорусов еще есть 5.5% I1, 5.5% R1b, 4% E1b, 2.5% J2, 1.5% G, 1% J1, 1% I2b, но ни одна из них не является характерно славянской. I1 – древнеевропейская гаплогруппа, или гаплогруппа Старой Европы (до III тыс. до н.э.), почти исчезнувшая в III тыс. до н.э., прошедшая «бутылочное горлышко популяции» и возродившаяся во II тыс. до н.э., и ставшая общеевропейской, как было описано выше; R1b – наследие ямной культуры и отчасти сарматов, потомков того же древнего рода R1b, прошедшего миграциями от Сибири до Днепра и далее через Кавказ в Месопотамию и затем в Европу разными миграционными путями, основной из них через Пиренеи и далее в континентальную Европу как культура колоколовидных кубков; E1b – потомки древних египтян и затем древних греков, расселившиеся по Средиземноморью и черноморским берегам; J2 – потомки древних бедуинов, составившие затем в заметной части население Средиземноморья и значительную часть евреев и арабов; J1 – преобладающая гаплогруппа у евреев и арабов, а также потомков древних евреев по всему миру, включая, например, горских евреев на Кавказе и значительную часть евреев-ашкенази, передвинувшихся в средние века (в особенности, после эпидемии черной чумы середины XIV века) во все славянские страны и нашедших там убежища от преследований; G – древнеевропейская гаплогруппа, еще одна из основных гаплогрупп Старой Европы, практически исчезнувшая, остатки которой бежали в Малую Азию, на Кавказ и в Переднюю Азию, и сейчас одна из основных кавказских гаплогрупп (например, до 75% у осетин); I1b – минорная гаплогруппа по всему миру, в Европе нигде не превышает 4-7% (в основном, в Северной Европе – Бельгия, Нидерланды, Дания, Норвегия, Швеция, северная Франция, северная и западная Германия), возможно, тоже реликт Старой Европы.
Примерно такие же вкрапления указанных минорых гаплогрупп явно неславянского происхождения есть по всем странам, включая славянские. Итак, перейдем к славянским странам, добавив балтов – литовцев и латышей, а также «северных русских», как, например, коми и марийцы, которые славянами, строго говоря, не являются, но которых популяционные генетики часто используют в качестве «этнических русских», и более того, заявляют их как эталон этнических русских, чтобы поднять в последних долю «финно-угорской компоненты». Именно в таком виде российские популяционные генетики Е. и О. Балановские заявили «геномный эталон этнических русских» в международную геномную базу данных, подробнее см. здесь. Так русские оказались в базе данных «финно-уграми».
Помимо этого, добавим в список гаплогруппу R1b, для славян нехарактерную, и которая унаследована в России от ямной культуры (найдена в составе ископаемых ДНК с археологическими датировками 2700-3300 гг. до н.э., Haak et al, 2015), отчасти от намного более поздних сарматов, в Европе – от потомков культуры колоколовидных кубков (позже 4800 лет назад).
Если не уходить в детали, то картина в целом ясна – у славян преобладает гаплогруппа R1a (за исключением балканских славян – болгар, сербов, хорват, македонцев, боснийцев, у которых доминирует I2a), за ней следует характерная южнославянская гаплогруппа I2a, и замыкает южно-балтийская гаплогруппа N1c1, которая к северо-востоку становится финно-угорской. Отдельно стоят балты – литовцы и латыши, у которых содержание R1a и N1c1 практически одинаково. Как мы видим, корни славян в отношении их ДНК-родового происхождения уходят в разные стороны.
Эти данные уже позволяют полностью предсказать, что покажут громоздкие данные «генетики», полученные по миллионам точек, и обработанные крайне формализовано. Результат же заранее известен. Ясно, что поляки, белорусы, русские и украинцы, то есть «восточные и западные славяне», у которых доминирует гаплогруппа R1a, образуют один «кластер» точек, в котором русские с более заметным вкладом гаплогруппы N1c1 будут несколько смещаться в сторону балтов (литовцев и латышей), еще больше туда будут смещаться «северные русские», которые уже смыкаются с финно-уграми. Напротив, сербы, хорваты, македонцы и боснийцы, то есть «южные славяне», у которых доминирует гаплогруппа I2a, образуют отдельный «кластер», а чехи и словаки, у которых, в отличие от других, повышено содержание гаплогруппы R1b (22% и 15%, соответственно), характерной для Западной и Центральной Европы, тяготеют к немцам, с еще более высоким содержанием этой гаплогруппы, причем чехи (с более высоким содержанием R1b) тяготеют больше, чем словаки.
Именно так и получилось в статье Кушнаревич и Балановских с соавторами. Цитируем:
>> «Восточные славяне – русские из центральных и южных регионов, белорусы и украинцы – образуют свой кластер»,
>> «напротив, русские из северных областей европейской части России отходят от остальных восточных славян, и располагаются вблизи своих финно-говорящих соседей»;
>> «чехи, и в меньшей степени словаки, сдвинуты по направлению к немцам и другим западноевропейцам»;
>> «поляки или совпадают с восточными славянами, или находятся близко к ним»;
>> «большинство южных славян… балканского полуострова… отделены от остальных балто-славянских популяций, и образуют размытую группу популяций, которые разделяются на западные регионы (словенцы, хорваты и боснийцы) и восточные регионы (македонцы и болгары)».
Действительно, смотрим на состав гаплогрупп македонцев и болгар – он практически идентичен по всем четырем основным гаплогруппам, приведенным выше.
>> «латыши и литовцы, говорящие на балтских языках, расположены вблизи эстонцев, говорящих на финно-угорских языках, если рассматривать Y-хромосому, и слегка сдвинуты к восточным славянам по аутосомам».
Тоже ясно, почему, у них поровну R1a (доминирующей у восточных славян) и N1c1 (доминирующей у финно-угров). Аутосомы добавляют «женский вклад», как видно, совсем незначительный, тем более в контексте обсуждаемой работы.
>> «мордвины показывают близость к обеим балтским популяциям, видимо, отражая исторические свидетельства, что племена, говорившие на балтских языках, расширялись на восток Восточно-Европейской равнины».
Первое само собой разумеется, поскольку у мордвин в среднем заниженное значение R1a (36%), по сравнению с большинством восточных славян и почти равное таковому у литовцев и латышей (38% и 40%) при почти том же значении N1c1 (15%), что у этнических русских. А второе – вообще неверно, это не балты продвигались на восток, а, напротив, носители гаплогруппы N1c1 продвигались на запад, и на этом пути достигли Балтики, что проявляется в составе гаплогрупп литовцев и латышей.
Так что «генетика» в данном исследовании ничего нового не привнесла в то, что давно известно по спискам опубликованных распределений гаплогрупп в славянских и соседних популяциях. И какой же вывод делают авторы из того, что давно известно? Вывода три (цитаты):
>> «начальное разделение протославян осталось невыясненным»,
>> «по лексикостатистическим данным разделение на западных, восточных и южных славян произошло примерно 1900 лет назад, дальнейшая диверсификация – примерно 1300-1500 лет назад, и расхождение на отдельные языки – 1000-500 лет назад».
>> «Y-хромосомная диверсификация на низшем уровне 1 популяционной структуры – среди локальных популяций, говорящих на одних и тех же языках – варьируется от почти 0 для чехов и македонцев до 0.05 в пределах северных русских, что в среднем дает 0.01. Генетическая дифференциация среди этнических популяций, относящихся к уровню 2 лингвистической ветви, приблизительно равна 0.03, и вариация в пределах ветви уровня 3 балто-славянских популяций увеличивается до 0.06».
Всё. На этом раздел статьи заканчивается. В этом состоит, по словам О. Балановского, то, что «в ближайшее десятилетие вряд ли можно ожидать чего-то принципиально нового в области генетики и лингвистики славянских популяций».
Видимо, этот «приципиально новый вклад» состоит, как полагают авторы обсуждаемой статьи, в том, что имеет место «корреляция» между деревом языков, приведенных в начале раздела, с географией, то есть «южными славянами», «северными», «западными» и «восточными». А состав гаплогрупп тоже определяется географией, поэтому было бы более чем странно, если бы такой корреляции не было. В итоге авторы статьи используют «циклический аргумент», когда территория определяет состав гаплогрупп, она же определяет разделение языков на локальные, а значит, состав гаплогрупп определяет разделение языков на локальные. Видимо, это и есть «принципиально новое».
Вот в описанном выше и состоит главное отличие популяционной генетики от ДНК-генеалогии. И дело не только в принципиально разной методологии, дело в принципах постановки и решения задач.
На обсуждении «данных лексикостатистики» можно и не останавливаться, ничего нового там нет, обработка данных крайне формализованная и невнятная. Да и в обсуждении филолингвистического дерева, приведенного выше, у авторов нет ничего нового. Они ссылаются на дерево Gray and Atkinson (2003):
Фрагмент дерева, относящийся к балто-славянским языкам (из статьи Gray and Atkinson, 2003)
но различия (или сходства) даже не обсуждают. Что говорить, когда нечего говорить? Лексикостатистика – вообще методология, мало кем в науке принятая, из-за обилия произвольных приближений и допущений, и авторы это еще раз показывают. В любом случае, никакой новой связи с генетикой не обнаружено, а то, что описано в статье – тривиально. Но давайте коротко остановимся на показанных деревьях, на данных лексикостатистики, и данных ДНК-генеалогии по балтам и славянам. О роли лексикостатистики в исследованиях, помимо замечания выше, можно сказать, что ей, как правило, отдается весьма низкий ранг достоверности, и ее данные могут служить только как вспомогательные, как дополнительное свидетельство в пользу того, что уже получено другими методами. Как мы видим, у Грея и Аткинсона (2003) расхождение славянских и балтских языков (литовского и латышского) датируется 3400 лет назад, то есть серединой II тыс. до н.э. То же воспроизводит и дерево в статье Кушнаревич и Балановских с соавторами (2015).
Это – крайне маловероятно и не стыкуется с данными ДНК-генеалогии, согласно которым в те времена будущие балты были еще на Урале или только начинали миграции на запад от Урала. Разумеется, никаких индоевропейских языков у них тогда не было. Были уральские, угорские или те, как это решили назвать лингвисты. К балтийскому побережью предки литовцев и латышей гаплогруппы N1c1 подойдут только к середине I тыс. до н.э. Датировки по 67-маркерным гаплотипам литовцев и латышей гаплогруппы N1c1 показывают время жизни их общих предков 2600±300 и 2100±300 лет назад, соответственно. Общий предок современных этнических русских гаплогруппы N1c1 жил 3300±340 лет назад, но это включает более древние зауральские и уральские гаплотипы, поэтому и общий предок несколько древнее. В Литве и Латвии такие пока не обнаружены.
Поэтому никакого «балто-славянского единства» или «единства гаплогрупп N1c1 и R1a» в середине II тыс. до н.э., и тем более ранее, просто не могло быть, как и «славянских языков, выделившихся из балтских». Ситуация была обратной – прибывшие на берега Балтийского моря предки литовцев и латышей, видимо, относительно малочисленные носители гаплогрупп N1c1 переняли ИЕ языки автохтонных (видимо, можно так назвать) носителей гаплогруппы R1a, которые обитали там как минимум тысячу лет (а если учесть ископаемую гаплогруппу R1a на Онежском озере с датировкой 7500 лет назад, в середине VI тыс. до н.э., то обитавшие к тому времени и 5 тысяч лет). Они и образовали «единство R1a и N1c1», которое с тех пор не распадалось, поскольку и сейчас у литовцев и латышей этих гаплогрупп поровну, примерно по 40% от всех. Языки разошлись, и, как будет показано ниже из данных С.А. Старостина, русский и литовский разошлись только в первой половине нашей эры.
В связи с этим с некоторым изумлением воспринимается основное положение лингвистов, специалистов по балтским языкам, что «балто-славянское единство иногда считают продуктом позднейшего сближения двух независимых индоевропейских диалектов», ссылка. Аналогично, давно затвержденное положение об особой индоевропейской архаичности литовского языка надо понимать, что архаичным был язык носителей гаплогруппы R1a, у которых предки литовцев этот язык переняли.
Вывод – славянские языки (и их предки) не «выделились» из балтских, а, напротив, были источником ИЕ языков для предков литовцев и латышей второй половины I тыс. до н.э. – первой половины I тыс. н.э.
Если мы обратим внимание на лексикостатистику в том варианте, с которым работал С.А. Старостин, то она, на первый взгляд, дает разумные результаты. Например, согласно его данным (Старостин, 1989), в современном русском и персидском языках в стословнике Сводеша имеется 28% совпадений. Это дает
от времени расхождения языков (до настоящего времени), что примерно соответствует времени ухода ариев на Иранское плато (0.05 – это по Старостину константа скорости выпадания слов из стословника ИЕ языков). Для русского и древнеиндийского языка имеется 54% совпадений, что соответствует
от времени расхождения языков. Это примерно равно времени, когда Панини создал классический санскрит. Какова бы ни была надежность этих интерпретаций, данные намного отличаются от степени расхождения русского и литовского языков, чему С.А. Старостин относил величину 74-76% (по разным текстам). Это соответствует времени расхождения 1660-1740 лет назад, то есть III-IV вв. нашей эры. Это никак не стыкуется с лексикостатистическими оценками ни Грея с Аткинсоном, ни Кушнаревич с сотрудниками. Не может три четверти совпадения слов из стословника соответствовать расхождению языков, начиная со столь давнего времени, как середина II тыс. до н.э.
Таким образом, мы видим, что подгонка данных «генетики» под заранее заданную, и, скорее всего, неверную датировку не только грубо нарушает принцип, сформулированный В.В. Седовым, но и ведет к неверным результатам как лингвистики, так и генетики. Основная ирония в том, что провозглашенная Балановским максима «в ближайшее десятилетие вряд ли можно ожидать чего-то принципиально нового в области генетики и лингвистики славянских популяций» оказалась опровергнута уже через две недели после этого провозглашения.
Клёсов А.А. Действительно ли «генетики нашли разных русских»?
Седов В.В. Происхождение и ранняя история славян. М., 1979.
Старостин С.А. Сравнительно-историческое языкознание и лексикостатистика. Лингвистическая реконст-рукция и древнейшая история Востока. М., 1989. С. 3-39.
Gray R.D., Atkinson Q.D. Language-tree divergence times support the Anatolian theory of Indo-European origin. Nature, 2003. 426, 435-439.
Kushniarevich, A., Utevska, O., Chuhryaeva, M., Agdzhoyan, A., Dibirova, K., Uktveryte, I., Möls, M., Mulahasanovic, L., Pshenichnov, A., Frolova, S., Shanko, A., Metspalu1, E., Reidla, M., Tambets, K., Tamm, E., Koshel, S., Zaporozhchenko, V., Atramentova, L., Kučinskas, V., Davydenko, O., Goncharova, O., Evseeva, I., Churnosov, M., Pocheshchova, E., Yunusbayev, B., Khusnutdinova, E., Marjanović, D., Rudan, P., Rootsi, S., Yankovsky, N., Endicott, P., Kassian, A., Dybo, A., The Genographic Consortium, Tyler-Smith, C., Balanovska, E., Metspalu1, M., Kivisild, T., Villems, R., Balanovsky, O. (2015) PLOS One, September 2, DOI: 10.1371/journal.pone.0135820
Анатолий А. Клёсов,
доктор химических наук, профессор