1 Институт Цитологии и Генетики, Сибирская Ветвь Российской Академии Наук, 10 Лаврентиев Авеню., Новосибирск, 630090, Россия; 2Институт Археологии и Этнографии, Сибирская Ветвь Российской Академии Наук 3Институт Терапии, Сибирская Ветвь Российской Академии Медицинских Наук + Автор-Корреспондент | ||||||||
Foreword | ||||||||
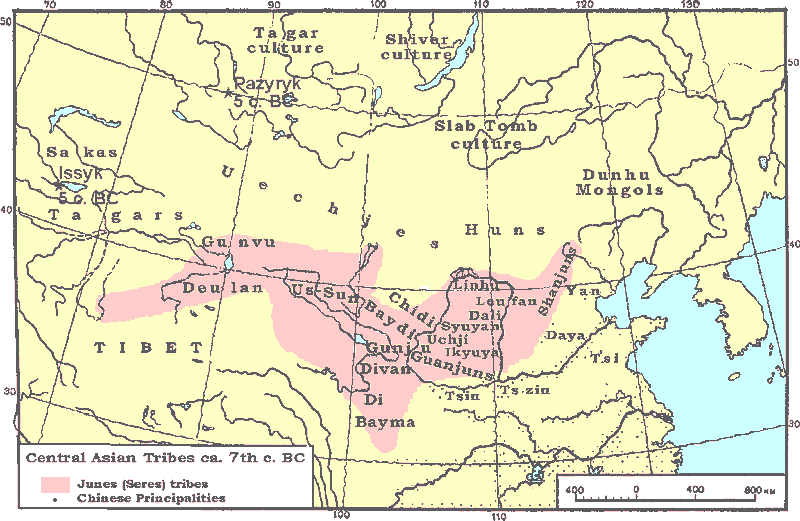
Pazyryk culture From Wikipedia, the free encyclopedia.
The Pazyryk culture (ca. 5th century BC) refers to an archaeological culture identified by excavated artefacts and mummified humans in the Siberian permafrost. The mummies are buried in kurgans similar to the kurgans of western Scythian culture in modern Ukraine. Archaeologists associate the sites with the widespread Scythian culture of the steppe. The vessels excavated from the tombs are similar to ones still in use today by indigenous groups.
Altaic peoples still profess beliefs that relies heavily on their kam's (aka shaman's) intellectual and spiritual knowledge, there is a belief that the disturbed sanctity are causing increased suicides, sickness and earthquakes in the region and hence the corps must be brought back to their origins to stop the chaos.
Ever since the first investigations in the 1920es, the Irano-enthusiasts labeled the Pazyryk people and their Scythian culture as invariably Iranian-lingual, building a variety of conjectures to fit the facts into their Indo-European schemes. Lately, however, the Iranian label tends to melt away, turning a glorious expression "Iranian-lingual Scythians" into a bleak and un-IE-patriotic simple "Scythians", and the its vestiges, scattered everywhere, and carrying very pronounced overtones, serve as a reminder of the past centuries' scientific anopia. Even now, in their scientific quests, investigators still manage to "get surprised" at every corner, discovering along the way that the eternal truths they were brought up with were conjured at odds with unpretentious reality.
The scientific investigation is in its baby steps only, we have a glimpse at mtDNA of the maternal lineages, as an aggregate value not resolved properly in time. Knowing that the moms as a group wore Paleosiberian genes is just a small step, we still want to know the composition of the pops, and their horses, and their accurate timing, and the bacteria used to make kumiss, and many many more.
| ||||||||
Links | ||||||||
| http://www.bionet.nsc.ru/bgrs/thesis/99/index.html | ||||||||
Summary | ||||||||
Keywords: molecular genetic techniques, human population genofond
| ||||||||
Структура контрольной области mtDNA выделенной из трех представителей Пазырыкской Культуры Горного Алтая (iv - ii столетие до н.э.), одной из центральных культур в Скифо - Сибирском мире, была проанализирована молекулярными генетическими методами.
В дополнение к техническим молекулярным биологическим проблемам, рассматривались антропогенные вопросы связанные с изученем специфичного материала:
(1), какая информация, определяющаяся етногенетического состава Пазырыка может быть получена от mtDNA экспертизы;
(2), что является относительной информативностью различных методических подходов к оценке полиморфизма;
(3), что является соответствием(корреспонденцией) между молекулярными генетическими и палеоантропологическими данными; и, наконец,
(4), чтобы определить роль Пазырыка в процессах развития человеческого генофонда населения в Евразии.
В случае женщины от могильного холма Ak-Alakha-5 (образец 1), ДНК была изолирована от ткани мышцы. В случае тканей в насыпях номер 1 и 3 из памятника Verkh-Kal'gzhin-II (образцы 2 и 3), использовалась легкая коричневатая масса, извлеченная из черепа и выделена макроскопически, как смесь жирных и мозговых тканей . Изоляция ДНК и удалениe PCR – ингибированного еффекта была сделана на отборной сорбции фрагментов ДНК на SiO2 частицах.
Затравки Thr (15898 - 5/-TCAAAGCTTACACCAGTCTTGTAAACC - 3/-15926) и TDKD (16518 - 5/-CCTGAAGTAGGAACCAGATG - 3/-16498) использовались для усиления контрольной области mtDNA.
Двунитевые продукты PCR получили используя образцы 1, и 2, клонированые в векторе PCRTMII (Invitrogene, TA клонирующий набор). ДНК плазмиды были получены порядки с использованием автоматизированного секвенатора и традиционно использованых коммерческих наборов (USB).
Первичная последовательность контрольной области mtDNA образца 3 была определена прямым секвенированием асимметричного продукта PCR; условия реакции были подобны тем где использовали для продукции двунитевого амплификатора с одним исключением: количество одной затравки было 50-кратно более низко чем другой.Каждый шаг изоляции ДНК сопровождался контрольными группами; все компоненты PCR смеси реакции были проанализированы, чтобы исключить любую возможность загрязнения с современной ДНК. Процессы были сделаны дважды с ДНК изолированной от двух образцов ткани для каждого индивидуума.
Первичная структура ДНК контрольной области mtDNA ~ 400 bp длиной была определена для каждого mtDNA образца, изолированного из трех Пазырыкцев. Показанные митотипы были сравнены с доступными mtDNA вариантами современных представителей Евразии.
Один переход впозиции 16,356 характерен для митотипа образца 1. Полностью идентичный вариант встречается только у Европоидов с частотой 1.2 % и распределен в них от Ближнего Востока до Исландии. Среди другого митотипа этой группы, 28.1 % отличается от митотипа 1 одной заменой (расположенный в различных позициях сравненного фрагмента); 36.5 %, двумя заменами; и 16.2, тремя заменами. Остальное митотипы показывает большее количество позиций с заменами. Фактически идентичные данные были получены для представителей Западно-сибирского Финно - Угорских народов (Khanty, Mansi, Komi, и Mari), занимая промежуточное положение между Европоидами и Монголоидами в их антропологических характеристиках. Из Монголоидов, вовлеченных в сравнение, варианты больше всего близко к митотипу 1 встречаются наиболее часто у Алтаицев, хотя они более редки чем у Европоидов. Группы митотипов с одной-тремя заменами у Монголоидов представлены 1, 5.9, и 5.9 %, соответственно, большая часть митотипов имеет больше чем три замены. Арктический Монголоид Дальнего Востока фактически не имеет подобных митотипов. Этот факт позволяет этому митотипу рассматриваться как наиболее определенный митотип для Европоидной mtDNA. Митотип 1 в его структуре соответствует хаплогрупе 1, показанной Richards и другими (1996) в митохондриальном генофонде Европейцев и занимающий там доминирующее положение.
Сравнительный анализ относительного подобия митотип 2 с вариантами, встречающимися в современных группах не сумел показывать очевидные различия в среднем количестве замен между представителями трех групп- Европоидами, Монголоидами, и промежуточных этнических групп. Фракция вариантов больше всего близких к митотипу 2 была относительно выше у групп Монголоидов, однако, оставаясь ниже чем фракция митотипа 1 наблюдаемый у Европоидов. Отмеченныме етноспецефические характеристики были найдены в последовательности митотипа 2. Три из этих четырех замен, отличающих этот митотип от Кембриджского митотипа (16223: С й Т; 16298: Т й С, и 16327: С й Т) соответствуют заменам, которые, вероятно, будут характерны для митотипs, принадлежащего хаплогрупе C, идентифицированой Уоллисом и Torroni в человеческом населении, базирующемся на полном рестрикционном анализе полного митохондриального генома. Эта хаплогрупа широко распространена среди местного населения Северной Азии и Америки, и ее частота уменьшена значительно в других областях. Высокая частота хаплогрупы C (~70 %) у Евенков была также подтверждена прямым секвенированием mtDNA образца представителей этой группы. Эти данные позволяют митотип 2 рассматривать как определенный Монголоидный вариант, встречающийся преобладающе в современных этнических группах Северной Азии (главным образом Евенков, Евен, и Селкуп народов) и к меньшему распространению в Центрально-азиатских группах.
Сравнительный анализ митотип 3 демонстрировал, что уровень средних различий между митотипами современных групп выше чем это для первых двух вариантов. Кроме того, отчетливо замечены более низкие количества в случае сравнения с Европоидами. Частота связанных вариантов также относительно низка, и они встречаются преобладающе в Европейских группах. Качественно, митотип 3 соответствует Европейской хаплогрупе 5. Данные доступные теперь предлагают, что этот хаплогрупа характеризована низкой частотой на всем протяжении Европы и встречаться в различных странах континента и в Ближнем Востоке. Было сообщено увеличение ее частоты у Финнов. Таким образом, выполненный анализ указывает, с одной стороны, относительно высокую специфичность митотипа 3 для современных Европейских групп и, с другой, его низкое наличие у всех исследованные образцов представителей групп современных Евроазийцев. Поскольку это было отмечено выше при обсуждении результатов рестрикционного анализа, открытие митотипа 3 по существу предлагает более высокую частоту хаплогрупы 5 митотипов в митохондриальном генофонде Пазырыка чем частота, наблюдаемая у современных этнических групп исследованных у Европоидов и Монголоидов.
Заключение
Специальный интерес было сравнение результатов молекулярного генетического исследования с современными палеоантропологическими характеристиками Пазырыкского населения.
Краниологическое исследование всех доступных материалов по погребениям этой культуры демонстрировали и Монголоидные и Европоидные компоненты в антропологическом составе Пазырыка. Краниологический вариант, встречающийся в скотоводческих племенах во II тысячелетии до н.э. на территориях Южного Таджикистана, Южной и Юго-западной Туркмении, и Северного Ирана представляют Европоидный компонент. В последующих эпохах, этот вариант исчезает с территории Западной и Середней Азии. Предполагалось, что носители этого морфологического комплекса были немногочисленны и постепенно ассимилировались в массу Восточно Средиземноморских Европоидов. Компонент Монголоида включает два антропологических типа. Один тип, автохтонный, был найден в Алтайском крае на границе Неолитического и Энеолитического периодов в людях, погребенных вНижнеТиткескенская и Каменная пещерах, и во второй половине II тысячелетия до н.э.в населении Каракол Культуры. Типичная комбинация антропологической характеристики параметров этих типов 1, подобных Южно-сибирскому комплексу, встреченному в настоящее время в Казахах, Киргизах, и некоторых группах Хакассов и Южно-Алтайских народов. Второй тип, Палеосибирский, доминирован на территории около Байкала в течение Неолитического периода. В настоящее время, он встречается только у Евенков в области Северного Байкала. Сравнение выше-описанных данных с результатами молекулярного генетического анализа демонстрирует, что в настоящее время невозможно дать всестороннюю интерпретацию всех аспектов антропогенеза Пазырыкских людей, связанного с палеоантропологическим соображением, главным образом вследствие ограничил молекулярных генетических данных относительно большинства современных групп Северных, Среднего, и Средней Азии. Однако, некоторые вопросы, в особенности вопрос, связанный с Палеосибирским компонентом, получили убедительное подтверждение и развитие. Данные, доступные в настоящее время также предлагают некоторые генетические связи Пазырыка с современным населением Горного Алтая. Открытие митотипа 2, обладающего отмеченным подобием с митотипами Северно-Азиатских Монголоидов, , в индивидууме с Европоидными антропологическими характеристиками - еще одно существенное открытие. Этот факт демонстрирует отсутствие абсолютного параллелизма в антропологических и молекулярных-генетических характеристиках. В заключении обратите внимание, что потенциал интерпретации полученных молекулярных генетических данных увеличится значительно с накоплением информации, для современных групп географически и исторически связанных Пазырыкскими людьми. http://www.turkicworld.org/ | ||||||||